- TGF-βがその受容体に結合すると、異なる細胞からさまざまな応答を引き出すことができる。どちらのタイプの細胞でもSmad3が活性化される。シグナル伝達経路が保存されているとすると、さまざまな細胞でTGF-βへの応答に多様性がある原因は何か。
- TGF-βの細胞表面受容体への結合によって生じたシグナルは、標的遺伝子の発現の起こる核へどのように伝えられるか。
- サイトカイン受容体と受容体型チロシンキナーゼに共通する三つの特徴を指摘せよ。これらの受容体の酵素活性に関する違い を一つ指摘せよ
- エリスロポエチンがその細胞表面受容体に結合したときに細胞内で進行する過程は、遺伝子発現を活性化する細胞のシグナル伝達経路の例として,よく調べられている。(a)JAK2がSTAT5を活性化したとき、および(b)GRB2がEpo受容体に結合したときに、細胞質から核へ移行する分子は何か。
- 活性化されたシグナル伝達経路が標的遺伝子の発現に適当な変化を与えた後には、その経路は不活性化されなければならない。さもないと、多数のがんにおいて増殖因子の経路がシグナルを送り続けているという例にみられるように、病理的な結果がもたらされかねない。多くのシグナル伝達経路は内在的に負のフィードバック機構をもっており、それによって一つの経路の下流で起こる過程が上流の過程を抑制する。(a) TGF-β,および(b) エリスロポエチンによって誘導されたシグナルを抑制的に調節する負のフィードバック機構を述べよ。
- GRB2は、それ自体は内在性の酵素活性をもたないにもかかわらず,MAP キナーゼを活性化する上皮増殖因子(EGF)のシグナル伝達経路にとって必須の構成成分である。GRB2の機能は 何か。GRB2の機能において,SH2およびSH3ドメインの果たす役割は何か。ほかにも多くのシグナル伝達タンパク質がSH2 ドメインをもっている。何がSH2と他の分子の相互作用の特異性を決めるのか。
- Ras タンパク質にある突然変異が起こると、それを恒常的に活性(RasD)にする。恒常的な活性化とは何か。恒常的に活性型のRasは、どのようにがんを進行させるか。どのようなタイプの突然変異が起こると、次のタンパク質が恒常的に活性型になるか。(a) Smad3 (b) MAP キナーゼ(c)NF-KB
- 出芽酵母 S. cerevisiae において,酵素Ste11はいくつかの異なるMAP キナーゼシグナル伝達経路に関与している。接合因子のシグナル伝達経路における Ste11の基質は何か。Ste11 は高浸透圧の培地によって誘導される MAP キナーゼシグナル伝達経路にも関与しているが、酵母細胞が接合因子によって刺激されたとき,何が高浸透圧下での生存に必要な菌系成長を妨げるのか。
- プロテインキナーゼBを完全に活性化するのに必要な過程を述べよ。筋細胞において、プロテインキナーゼBによって伝えられるインスリンの二つの効果を指摘せよ。
- PI 3-キナーゼシグナル伝達経路におけるPTEN ホスファターゼの機能を述べよ.PTEN の機能喪失型変異ががんを進行させるのはなぜか。恒常的に活性型であるPTEN の細胞増殖とその生存に与える影響を予測せよ。
- プロテインキナーゼ A (PKA)の活性化は、細胞に短期間と長期間の効果の両方を与えることができる。肝細胞において活性化されたPKAによるいくつかの長期間の効果は何か。それらの効果はいかなる経路によって仲介されるか。
- Wnt タンパク質と同じように、細胞外シグナル伝達タンパク質のHedgehog は細胞膜につなぎとめられている。Hedgehogの膜結合性は、その分子へのいかなる修飾によるか。この特性は、なぜ有用か。
- 鞭毛内輸送(IFT)タンパク質は、繊毛や鞭毛内での物質輸送に必要である。多くの細胞は、一次繊毛とよばれる単一の動かない繊毛をもっている。この一次繊毛がシグナル伝達に介在する証拠は何か。
- NF-κBを活性化するシグナル伝達経路が、サイトカインあるいは受容体型チロシンキナーゼのシグナル伝達経路に比べて相対的に不可逆的だと考えられるのはなぜか。それにもかかわらず、NF-κBは結果としては抑制的な調節を受けなければならない。NF-κBシグナル伝達経路のスイッチはどのようにして切られるか。
- Yーセクレターゼによって触媒される生化学反応は何か。Deltaの Notch 受容体への結合で誘導されたシグナルを伝達する際に、ソーセクレターゼの果たす役割は何か。ソーセクレターゼのサブユニットの一つであるプレセニリン1の突然変異は、アルツハイマー病にどう寄与すると考えられているか。
TGF-βがその受容体に結合すると、異なる細胞からさまざまな応答を引き出すことができる。どちらのタイプの細胞でもSmad3が活性化される。シグナル伝達経路が保存されているとすると、さまざまな細胞でTGF-βへの応答に多様性がある原因は何か。
TGF-βは様々な細胞で共通のシグナル伝達(Smad3の活性化など)を利用しますが、細胞ごとのTGF-β応答が多様になる主な原因は各細胞における「補助因子・転写調節タンパク質」「エピジェネティック状態」「他シグナル経路とのクロストーク」などが異なるためと考えられます。
多様性の要因
- 補因子(転写調節因子)の違い
- エピジェネティック状態の違い
- 他のシグナル経路とのクロストーク
これらの理由から、同じTGF-β刺激でも上皮細胞ではプラスミノーゲンアクチベーターインヒビター、B細胞では免疫グロブリン(IgAなど)と応答が異なり、細胞の種類ごとにTGF-β応答の多様性が生み出されているのです。
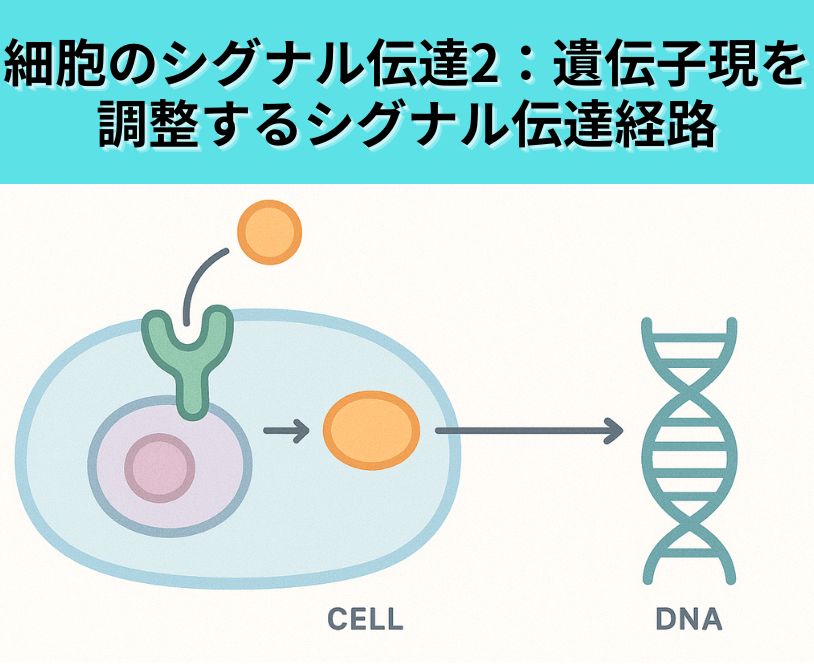
TGF-βの細胞表面受容体への結合によって生じたシグナルは、標的遺伝子の発現の起こる核へどのように伝えられるか。
- TGF-βが細胞表面のII型受容体(TβRII)に結合すると、I型受容体(TβRI)と複合体を形成します。
- II型受容体のキナーゼ活性によってI型受容体のセリン/スレオニン残基がリン酸化され、I型受容体が活性化されます。
- 活性化されたI型受容体は、細胞質のSmad2・Smad3タンパク質のC末端をリン酸化します。
- リン酸化されたSmad2/3はSmad4(共通Smad)と結合し、複合体を形成します。
- このSmad複合体は核内へ移行し、標的遺伝子のプロモーター領域で他の転写因子や補助因子と協調して転写制御を行い、標的遺伝子の発現を誘導します。
このように、TGF-β受容体で始まったシグナルはSmadタンパク質を介して核へ運ばれ、標的遺伝子発現の制御まで伝達されます。Smad以外にもMAPKなど非定型経路を通る場合がありますが、基本的な核転送は上記のSmad経路が中心です。
サイトカイン受容体と受容体型チロシンキナーゼに共通する三つの特徴を指摘せよ。これらの受容体の酵素活性に関する違い を一つ指摘せよ
サイトカイン受容体と受容体型チロシンキナーゼに共通する三つの特徴
- 細胞表面に存在する膜貫通タンパク質である
両者とも細胞外にリガンド結合部位をもち、膜を貫通する構造でシグナルを細胞内に伝える。 - リガンド結合による二量体化またはオリゴマー化が起こる
リガンドが結合すると、受容体同士が集合し、細胞内の活性化のきっかけとなる。 - 細胞内にシグナル伝達のためのチロシンリン酸化部位を持つ
リガンド刺激で細胞内の特定のチロシン残基がリン酸化され、下流分子の結合や活性化を促す。
酵素活性に関する違い
- 受容体型チロシンキナーゼ(RTK)は受容体自身がチロシンキナーゼ活性を持ち、自己リン酸化が起こるのに対し、
- サイトカイン受容体は受容体自体にチロシンキナーゼ活性がなく、JAK(非受容体型チロシンキナーゼ)が受容体に結合してその活性化とリン酸化を行う点に違いがあります。
この違いにより、RTKは自身のキナーゼドメインで直接シグナル開始を行う一方で、サイトカイン受容体はJAKとの相互作用を介して間接的にシグナルを伝達します。
エリスロポエチンがその細胞表面受容体に結合したときに細胞内で進行する過程は、遺伝子発現を活性化する細胞のシグナル伝達経路の例として,よく調べられている。(a)JAK2がSTAT5を活性化したとき、および(b)GRB2がEpo受容体に結合したときに、細胞質から核へ移行する分子は何か。
(a) JAK2がSTAT5を活性化したときに核へ移行する分子
- STAT5(Signal Transducer and Activator of Transcription 5)がリン酸化されて活性化されると、二量体化し細胞核へ移行します。活性化したSTAT5は核内で標的遺伝子の転写を促進します。
(b) GRB2がEpo受容体に結合したときに核へ移行する分子
- GRB2はシグナル伝達アダプタータンパク質であり、Epo受容体に結合してRas-MAPKカスケードを活性化します。
- この経路の下流で活性化される転写因子の一例にElk-1があり、Elk-1がリン酸化されて核へ移行し転写活性化を行います。
したがって
- (a)では、活性化されたSTAT5二量体が核へ移行し、
- (b)では、リン酸化され核へ移行する転写因子Elk-1などが主要な核内移行分子となります。
活性化されたシグナル伝達経路が標的遺伝子の発現に適当な変化を与えた後には、その経路は不活性化されなければならない。さもないと、多数のがんにおいて増殖因子の経路がシグナルを送り続けているという例にみられるように、病理的な結果がもたらされかねない。多くのシグナル伝達経路は内在的に負のフィードバック機構をもっており、それによって一つの経路の下流で起こる過程が上流の過程を抑制する。(a) TGF-β,および(b) エリスロポエチンによって誘導されたシグナルを抑制的に調節する負のフィードバック機構を述べよ。
(a) TGF-βの負のフィードバック機構
- TGF-βシグナル伝達を抑制する代表的な負のフィードバック因子としてSmad7があり、TGF-β刺激により細胞内で発現誘導されます。
- Smad7はTGF-β受容体の細胞内ドメインに結合し、R-Smad(Smad2/3)のリン酸化を阻害してシグナルの伝達を抑制します。
- また、Smad7の他にTMEPAIファミリーなども負の調節に関与しています。これらはTGF-βシグナルの過剰活性化を防ぐ役割を果たし、シグナルの適切な制御を担います。
(b) エリスロポエチンの負のフィードバック機構
- エリスロポエチン(Epo)シグナル伝達の負のフィードバックには主に**SOCSファミリー(Suppressor of Cytokine Signaling)**が関与します。
- SOCSタンパク質はEpo受容体やJAK2キナーゼに結合し、それらのリン酸化を妨げることでシグナルの抑制を行います。
- これにより、Epo経路の過剰活性化を防ぎ、シグナルの適切な終了や細胞応答の調整を行う重要な負のフィードバック機構となっています。
以上のように、TGF-βとエリスロポエチンはいずれも自らのシグナル伝達経路の下流で負のフィードバック調節因子を誘導し、シグナル過剰活性過剰活性化を防いで細胞の恒常性を維持しています。
GRB2は、それ自体は内在性の酵素活性をもたないにもかかわらず,MAP キナーゼを活性化する上皮増殖因子(EGF)のシグナル伝達経路にとって必須の構成成分である。GRB2の機能は 何か。GRB2の機能において,SH2およびSH3ドメインの果たす役割は何か。ほかにも多くのシグナル伝達タンパク質がSH2 ドメインをもっている。何がSH2と他の分子の相互作用の特異性を決めるのか。
GRB2の機能
- GRB2はアダプタータンパク質としての役割を持ちます。自身に酵素活性は持たないものの、受容体や他タンパク質と結合してシグナル伝達経路の架け橋(スカフォールド)となり、下流の効果器タンパク質をリクルートして活性化を促進します。
- 具体的には、EGF受容体が活性化されてリン酸化されると、GRB2がそのリン酸化チロシンに結合し、下流のGEF(グアニンヌクレオチド交換因子)Sosを引き寄せます。SosはRasを活性化し、MAPK経路の活性化へとつながります。
SH2ドメインとSH3ドメインの役割
- SH2ドメイン
リン酸化されたチロシン残基を含む特定のアミノ酸配列(pY-X-N-X motif)に特異的に結合し、リン酸化受容体や他のタンパク質を認識してGRB2をリクルートします。この相互作用がシグナルの開始点となります。 - SH3ドメイン
プロリンリッチ領域を持つタンパク質(例えば、Sosのプロリンリッチ領域)に結合し、下流シグナル伝達タンパク質同士の連結や活性化を媒介します。
SH2ドメインの結合特異性
- SH2ドメインと他タンパク質の相互作用の特異性は、**リン酸化チロシン周辺のアミノ酸配列(特にpY-X-N-Xなどのモチーフ)**により決まります。
- その微妙な配列の違いが、SH2ドメインの結合親和性や特異性を生じさせ、細胞内でのシグナル経路の選択と制御を可能にしています。
GRB2はSH2ドメインにより受容体リン酸化部位に結合し、SH3ドメインによりSosをリクルートしてMAPK経路の活性化を媒介する重要なアダプタータンパク質です。SH2ドメインの特異性はリン酸化チロシン周辺の配列によって制御されています。
Ras タンパク質にある突然変異が起こると、それを恒常的に活性(RasD)にする。恒常的な活性化とは何か。恒常的に活性型のRasは、どのようにがんを進行させるか。どのようなタイプの突然変異が起こると、次のタンパク質が恒常的に活性型になるか。(a) Smad3 (b) MAP キナーゼ(c)NF-KB
恒常的な活性化とは
- **恒常的活性化(コンスティテューティブアクティベーション)**とは、通常リガンドなどの刺激を必要とするシグナル分子が、突然変異の結果、常に活性化された状態にあり、シグナル伝達を継続的に行う状態を指します。
- Rasの場合、GTPの加水分解(GTPase活性)が低下したり失われる突然変異(特にコドン12, 13, 61の変異)により、RasがGDP型の不活性状態に戻れず活性型のまま持続的にシグナルを送り続けます。
恒常的活性型Rasががんを進行させる仕組み
- 活性化されたRasは、MAPキナーゼ経路(RAF→MEK→ERK)やPI3K/AKT経路などの下流シグナルを常に活性化します。
- これが細胞増殖、生存、回避機構の持続的刺激となり、細胞の制御性増殖、形質転換、腫瘍形成を引き起こし、がんの発生と進展に寄与します。
突然変異によって恒常的に活性化するタイプ
- (a) Smad3
通常は受容体キナーゼによるリン酸化によって活性化されるが、突然変異による恒常活性化はめずらしい。代わりに核内局在異常や補因子変化により活性調節異常を起こす場合がある。 - (b) MAPキナーゼ(ERKなど)
MAPK自体が活性型となる突然変異はまれで、多くはRasやRAFの変異により下流で恒常的に活性化される。 - (c) NF-κB
NF-κBは多くの細胞ストレスや炎症シグナルで活性化される転写因子で、IκBの分解異常や上流シグナルの変異により恒常的活性化される例がある。
要約すると、Rasの点突然変異による恒常的活性化はがん化の主要因であり、MAPK経路はこれにより持続活性化される。Smad3の変異で恒常活性化される例は稀であり、NF-κBはIκBやシグナル経路異常で活性化が持続しますす。
出芽酵母 S. cerevisiae において,酵素Ste11はいくつかの異なるMAP キナーゼシグナル伝達経路に関与している。接合因子のシグナル伝達経路における Ste11の基質は何か。Ste11 は高浸透圧の培地によって誘導される MAP キナーゼシグナル伝達経路にも関与しているが、酵母細胞が接合因子によって刺激されたとき,何が高浸透圧下での生存に必要な菌系成長を妨げるのか。
(1) 接合因子シグナル伝達でのSte11の基質
- Ste11はMAPキナーゼキナーゼキナーゼ(MAPKKK)として機能し、接合因子(フェロモン)シグナル伝達経路において、MAPキナーゼキナーゼ(MAPKK)であるSte7を基質としてリン酸化・活性化します。
- 活性化されたSte7は下流のMAPキナーゼ(Fus3など)を活性化し、接合因子応答を駆動します。
(2) 高浸透圧誘導MAPキナーゼ経路とSte11の関与
- Ste11は接合因子応答だけでなく、高浸透圧条件下で活性化されるHOG(High Osmolarity Glycerol)経路にも関与します。
- しかし、接合因子によって細胞が刺激された際、HOG経路の活性化および関連するシグナルが、高浸透圧下で菌糸様成長を妨げる要因となります。
- 高浸透圧条件では、HOG経路のMAPKKK Bck1およびSte11が、ストレス応答として細胞壁の改変や適応機構を活性化し、その結果、接合因子誘導の菌系成長が抑制されます。
まとめると、Ste11は接合因子シグナルではSte7をリン酸化し活性化する基質として働き、高浸透圧条件下ではHOG経路に関わり、細胞のストレス応答を活性化することが菌系成長の妨げとなります。
プロテインキナーゼBを完全に活性化するのに必要な過程を述べよ。筋細胞において、プロテインキナーゼBによって伝えられるインスリンの二つの効果を指摘せよ。
プロテインキナーゼB(Akt)の完全活性化過程
- PI3Kの活性化
インスリンや成長因子のシグナルにより、PI3キナーゼ(PI3K)が活性化され、PIP2(ホスファチジルイノシトール-4,5-二リン酸)をリン酸化してPIP3(ホスファチジルイノシトール-3,4,5-三リン酸)を生成します。 - Aktの膜局在化とリン酸化
PIP3に結合したAktは膜に局在化し、次にPDK1(phosphoinositide-dependent kinase-1)によってThr308残基がリン酸化され、活性化される。 - mTORC2によるセリン473のリン酸化
AktはさらにmTORC2によってセリン473残基もリン酸化され、完全に活性化される。 - Aktの下流標的の調節
活性化されたAktは、様々な下流標的(Bcl-2ファミリー、GSK-3、TSC2、FOXOなど)をリン酸化し、細胞増殖、生存、代謝調節、アポトーシス抑制に働きます。
インスリンによる筋細胞の二つの主要な効果
- グルコース取り込みの促進
Aktはグルコース輸送タンパク質(GLUT4)を細胞膜に移動させ、血中のグルコースを細胞内に取り込みやすくします。これにより血糖値の低下を促進。 - グリコーゲン合成の促進
AktはGSK-3をリン酸化し、γ-グルコシル基転移酵素の活性を解除してグリコーゲン合成を促進します。
まとめ
インスリンによるPI3K-Akt経路の活性化とその後のリン酸化イベントにより、筋細胞では血糖コントロールとエネルギー貯蔵に重要な効果がもたらされます。
PI 3-キナーゼシグナル伝達経路におけるPTEN ホスファターゼの機能を述べよ.PTEN の機能喪失型変異ががんを進行させるのはなぜか。恒常的に活性型であるPTEN の細胞増殖とその生存に与える影響を予測せよ。
PTENホスファターゼの機能
- PTENは脂質ホスファターゼであり、細胞膜上のホスファチジルイノシトール3,4,5-三リン酸(PIP3)を脱リン酸化してPIP2に戻す働きを持つ。
- この反応は、PI3キナーゼによるPIP3生成の逆反応であり、PI3K/Aktシグナル伝達経路を負に制御し、細胞の増殖・生存・遊走を抑制する働きがある。
PTENの機能喪失型変異ががんを進行させる理由
- PTENが機能喪失すると、PIP3の分解が阻害されて細胞膜上のPIP3濃度が過剰に高まる。
- これによりAktが過剰に活性化され、細胞増殖や生存シグナルが持続的に送られるため、細胞の制御のきかない増殖やアポトーシス抵抗性が獲得され、がんの発生や進展を促進する。
- したがって、PTENは重要ながん抑制因子として認識されている。
恒常的に活性型であるPTENの細胞増殖や生存に与える影響
- 恒常的に機能するPTENは、過剰なPIP3蓄積を防ぎ、Aktや他の下流シグナルの活性を抑制するため、細胞増殖が抑制され、生存や代謝が制御される。
- これにより、細胞周期の進行が適切にコントロールされ、不要な細胞増殖やがん化のリスクが低減されると予想される。
- しかし活性が強すぎると、逆に細胞の成長低下やアポトーシス誘導が増加する可能性もある。
PTENはPI3K/Akt経路の負のレギュレーターであり、その機能喪失はシグナルの過剰活性化を通じて腫瘍形成に寄与します。一方で恒常的活性は細胞の増殖抑制と生存制御に資すると言えます。
プロテインキナーゼ A (PKA)の活性化は、細胞に短期間と長期間の効果の両方を与えることができる。肝細胞において活性化されたPKAによるいくつかの長期間の効果は何か。それらの効果はいかなる経路によって仲介されるか。
肝細胞におけるPKA活性化の長期間効果
- 遺伝子発現の調節
活性化したPKAは核内に移行し、転写因子(例:CREB:cAMP応答エレメント結合タンパク質)をリン酸化して活性化します。CREBが標的遺伝子のプロモーターに結合し、遺伝子発現を促進します。 - 代謝関連遺伝子の発現変化
肝細胞では、グルコース新生酵素や脂質代謝を調節する酵素の遺伝子発現が制御され、長期的な代謝調節が行われます。これにより糖代謝や脂質代謝が調整され、エネルギー恒常性の維持に寄与します。
経路の仲介
- PKA活性化→CREBリン酸化→遺伝子のcAMP応答エレメント(CRE)への結合→転写活性化
- この経路が長期的な細胞応答を仲介し、代謝酵素の合成や分解などを制御します。
このように、PKAの長期間効果は主にCREBなどの転写因子を介した遺伝子発現制御によって媒介され、肝細胞における代謝の恒常性維持と調節に重要な役割を果たします。
Wnt タンパク質と同じように、細胞外シグナル伝達タンパク質のHedgehog は細胞膜につなぎとめられている。Hedgehogの膜結合性は、その分子へのいかなる修飾によるか。この特性は、なぜ有用か。
Hedgehogの膜結合性をもたらす修飾
- Hedgehog(Hh)タンパク質は、成熟過程で脂質修飾を受けます。
- 具体的には、N末端にパルミトイル化(脂肪酸のパルミチン酸の付加)が、C末端にコレステロール修飾が行われます。
- これらの脂質修飾により、Hhは強固に細胞膜や細胞外マトリックスに結合します。
なぜこの膜結合性が有用か
- 脂質による膜結合はHhの細胞間拡散を制限し、局所的なシグナリングを可能にするため重要です。
- Hhは膜結合性が高いため、発生過程における細胞の近傍の標的細胞に対して限定的かつ精密にシグナルを伝達し、精密な組織パターン形成に寄与します。
- また、Hhが過剰に広がるのを防ぎ、シグナルの空間的・濃度勾配を形成することで、細胞運命の制御の精度を高めます。
このようにHedgehogの膜結合性は脂質修飾によって制御されており、その特性が効果的かつ限定的なシグナル伝達と組織形成の鍵となっています。
鞭毛内輸送(IFT)タンパク質は、繊毛や鞭毛内での物質輸送に必要である。多くの細胞は、一次繊毛とよばれる単一の動かない繊毛をもっている。この一次繊毛がシグナル伝達に介在する証拠は何か。
- 一次繊毛は多くの哺乳類細胞表面に存在する不動性の細胞小器官で、細胞外のシグナル受容体やイオンチャネルが高密度に局在していることが明らかになっています。これは一次繊毛が細胞外の化学的・機械的シグナルを受容し、細胞内に伝達するアンテナの役割を果たしていることを示します。
- HedgehogやWnt、BMPなどの重要な発生シグナル伝達経路が一次繊毛に依存しており、これらのシグナル伝達の異常が繊毛形成異常や発生異常、遺伝性疾患(繊毛病)に結びついていることが強い証拠です。
- さらに、一次繊毛の形成・短縮・消失が細胞周期やシグナル感受性の調節に影響を与えることも示され、一時的に一次繊毛の存在が細胞のシグナル伝達のオンオフを制御していることを示唆しています。
これらの点から、一次繊毛はIFTタンパク質によって維持され、細胞のシグナル伝達を効率的にかつ選択的に行うための重要な細胞構造として機能していると考えられています。
NF-κBを活性化するシグナル伝達経路が、サイトカインあるいは受容体型チロシンキナーゼのシグナル伝達経路に比べて相対的に不可逆的だと考えられるのはなぜか。それにもかかわらず、NF-κBは結果としては抑制的な調節を受けなければならない。NF-κBシグナル伝達経路のスイッチはどのようにして切られるか。
NF-κBシグナル伝達経路が相対的に不可逆的と考えられる理由
- NF-κB経路は通常細胞質でIκBという阻害タンパク質と結合し不活性状態にありますが、刺激によりIκBが速やかにユビキチン依存的に分解されることで、NF-κBが核へ移行し遺伝子発現を活性化します。
- このIκBの分解は不可逆的なタンパク質分解反応であり、他のシグナル経路のように単なるリン酸化や脱リン酸化だけで制御される可逆的変化とは異なります。
- したがって、一旦活性化すると信号のオンが外部刺激なしには自発的に戻りにくく、不可逆的に近い「決定的なスイッチ」の特徴をもつと考えられます。
NF-κBシグナル伝達経路が抑制的調節を受ける必要性
- NF-κBは炎症反応や細胞増殖を強力に誘導するため、過剰な活性化は自己免疫疾患や炎症性疾患、がんの進展に繋がります。
- そのため、細胞は過剰なNF-κB活性化を抑制するための厳密な負のフィードバック制御機構を備えています。
NF-κBシグナル伝達経路のスイッチの切り方
- 代表的には、NF-κBによる標的遺伝子の転写産物であるIκBαタンパク質が新たに合成され、核からNF-κBを細胞質に戻して再び不活性化させます(負の自己調節ループ)。
- さらに、A20やその他の負の制御タンパク質がIKK複合体や上流のシグナル伝達成分を阻害し、シグナルの過剰活性を抑制します。
- これらの負のフィードバックは、細胞のNF-κB活性を一過性に保ち、シグナル伝達の振動現象や強度制御に寄与します。
NF-κB経路はIκB分解の不可逆性によりスイッチ的な活性化機構を持つが、その強力な作用ゆえに新規IκBやA20などの負のフィードバックで活性を厳密に制御し、細胞の恒常性を保っています。
Yーセクレターゼによって触媒される生化学反応は何か。Deltaの Notch 受容体への結合で誘導されたシグナルを伝達する際に、ソーセクレターゼの果たす役割は何か。ソーセクレターゼのサブユニットの一つであるプレセニリン1の突然変異は、アルツハイマー病にどう寄与すると考えられているか。
γ-セクレターゼによって触媒される生化学反応
γ-セクレターゼは膜タンパク質の**膜内切断(イントラメンブレンプロテアーゼ活性)**を触媒する酵素複合体です。特に、アミロイド前駆体タンパク質(APP)を切断してアミロイドβペプチドを産生します。
DeltaのNotch受容体シグナル伝達におけるソーセクレターゼの役割
Notch受容体はDeltaとの結合により、まず細胞外ドメインが切断(S2切断)され、続いてソーセクレターゼによる膜内ドメインの切断(S3切断)が起こります。
この膜内ドメインが切り出されることで活性化されたNotchの細胞内ドメイン(NICD)が遊離し、核へ移行して標的遺伝子の転写を調節します。
プレセニリン1の突然変異とアルツハイマー病への寄与
- プレセニリン1はγ-セクレターゼ複合体の主要サブユニットであり、その変異はγ-セクレターゼの切断特異性を変化させます。
- 特にアミロイドβペプチドの長い型(Aβ42, Aβ43)が過剰に産生され、これらは不溶性で神経毒性が強く、脳に蓄積してアルツハイマー病の原因となる老人斑を形成します。
- そのため、プレセニリン1遺伝子の変異は家族性アルツハイマー病の主要な原因と考えられています。
γ-セクレターゼは膜タンパク質の膜内切断を行う酵素であり、Notchシグナルの活性化やAPPの切断に関与します。プレセニリン1変異はこの酵素の機能を変え、アルツハイマー病の病態形成に寄与します。
問題文引用元:東京化学同人 分子細胞生物学 第6版

コメント