- 脳や他の神経系でのグリア細胞の役割は何か
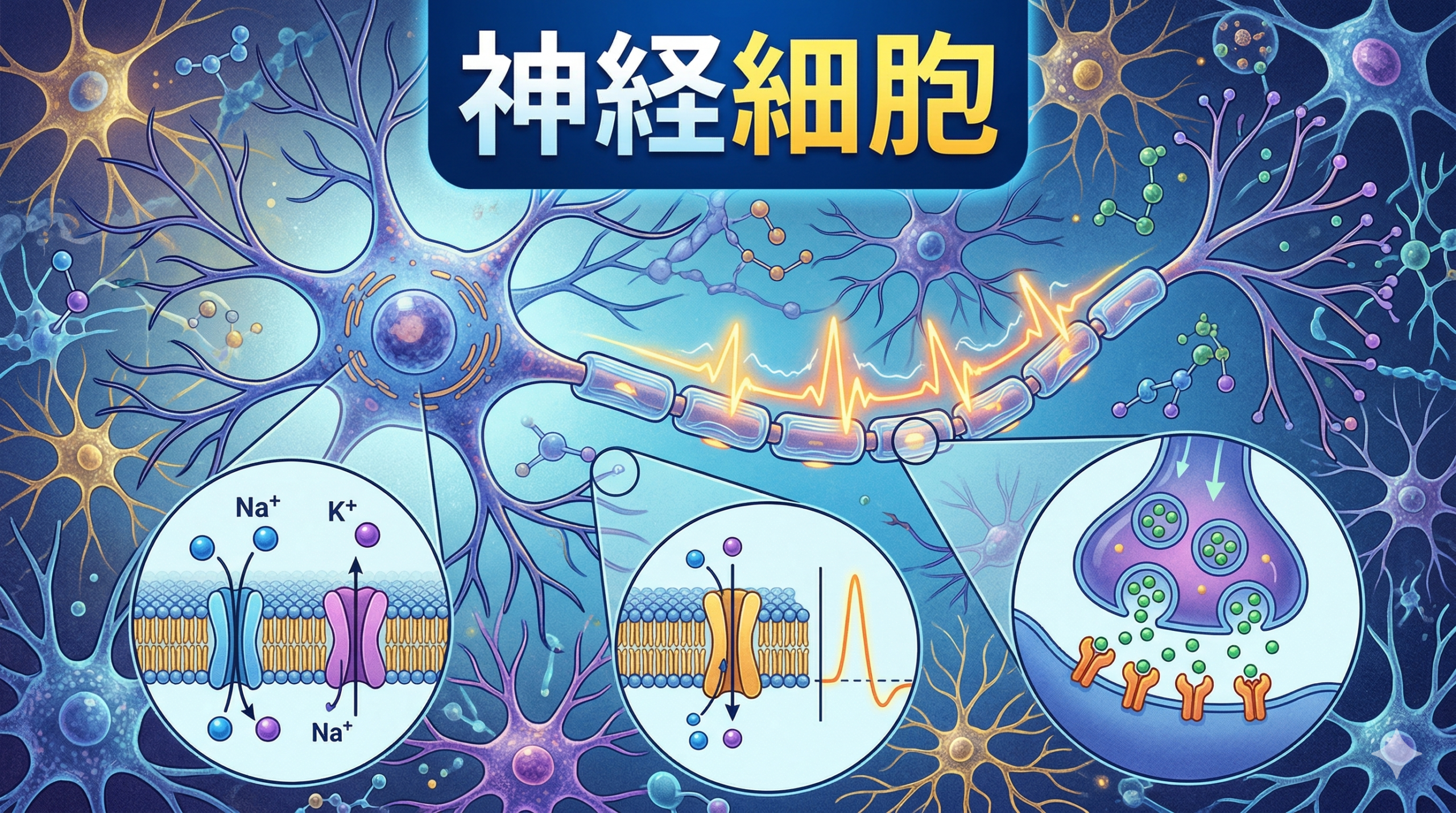
- 神経細胞の静止電位は細胞の外部と比べ内部が一60mVである。動物細胞ではこの静止電位はどのように維持されているか。
- 活動電位の三つの段階の名称を述べよ。それぞれの背景にある関連する分子基盤とイオンについて記述せよ。なぜ電位依存性チャネルという名称が活動電位の発生にかかわる Na+チャネルに使われているのか。
- K+チャネルの結晶構造から、電位感受性ドメインがこのタンパク質の他の部分とどのように相互作用して開閉していると考えられているか、説明せよ。この構造ー機能相関は他の電位依存性イオンチャネルではどのように適用できるか。
- なぜ活動電位の強度が軸索の末端へと伝わっていく際に下がらないのか説明せよ。
- 神経シグナルが細胞体に向かって逆行することを、何が防いでいるのか。
- ミエリン形成によって軸索に沿った活動電位の伝播速度が上がる。ミエリン形成とは何か。ミエリン形成によって、軸索に沿って存在するランビエ絞輪における電位依存性 Na+チャネルと Na+/K+ポンプが集中化されている。ランビエ絞輪間の間隔を10倍に増すと、活動電位の伝播にどのような影響が出ると予想されるか。
- アセチルコリンはシナプスで放出される一般的な神経伝達物質である。神経筋シナプスにおいてアセチルコリンエステラーゼの活性が低下した筋肉活動はどのような結果になるか、予測せよ。
- 刺激された細胞で活動電位がシナプスに到達すると、シナプス小胞は速やかにシナプス前細胞の細胞膜に融合する。これは1ミリ秒以内に起こる。このように驚くべき速さでこの現象が起こるのはどのような機構によるか。
- 脳においては特に、神経細胞は複数の興奮性と抑制性のシグナルを受け取る。神経細胞のこのシグナルを受け取る突起の名前は何か。神経細胞はこのシグナルをどのように統合し、活動電位を生じさせるか否かを決定しているか。
- シナプス小胞の再利用において、ダイナミンはどんな役割をしているか。それを支持する証拠は何か。
- 桿体細胞や錐体細胞が光によって刺激されたとき、一連の現象がカスケードとして実行され、それは最終的に視覚情報として脳へ伝達される。ロドプシンによる光吸収から桿体と錐体の細胞膜の過分極に至るこのカスケードの分子的な過程を概説せよ。
- 視覚におけるコントラスト(差異)の検出にはどのような細胞がかかわっているか。これらの細胞は、コントラストを検出するために光受容細胞の情報をどのように統合しているか。
- カエルにおける Roger Sperry の実験では、神経細胞がどのように接続されていくかを説明するための共鳴仮説と化学親和仮説をどのように区別したか。
脳や他の神経系でのグリア細胞の役割は何か
グリア細胞の基本機能
グリア細胞(glial cell、神経膠細胞)は、中枢神経系(脳・脊髄)でニューロン(神経細胞)を数で上回る支持細胞群です。物理的支持だけでなく、神経信号の調整、栄養供給、免疫防衛、修復など多岐にわたる役割を担います。
種類別役割の詳細
アストロサイト(星状膠細胞)
アストロサイトは星形の突起を持ち、ニューロンの周囲を密に包み込みます。
- 神経伝達物質の回収: ニューロンから放出されたグルタミン酸(興奮性神経伝達物質)やGABA(抑制性神経伝達物質)を速やかに取り込み、再利用します。これにより神経興奮の過剰(興奮毒性)を防ぎます。
- イオンバッファリング: 活動電位発生時に軸索外へ漏れ出るK⁺(カリウムイオン)を吸収・拡散し、神経信号の正確性を保ちます(空間的バッファリング)。
- 血液脳関門(BBB)の構築: 毛細血管内皮細胞と密着結合を形成し、血液由来の有害物質(毒素、重金属)を脳内に侵入させません。
- 代謝支持: グリコーゲンを貯蔵し、乳酸としてニューロンに供給。記憶形成時の高エネルギー需要に対応します。
- シナプス形成・可塑性の制御: シナプス(ニューロン間接合部)を物理的に包囲し、D-セリン(グリア由来神経伝達物質)を放出してNMDA受容体(学習に関わるイオンチャネル)を活性化。長期増強(LTP、記憶の基盤)に必須です。
オリゴデンドロサイト(少突起膠細胞)
オリゴデンドロサイトは少数の突起で多数の軸索を覆います。
- ミエリン鞘形成: 軸索を脂質豊富な多層膜で巻きつけ、電気信号の容量性漏れを防ぎます。これにより連続伝導が飛躍伝導(絶縁部分をスキップ)に変わり、伝達速度が100倍以上に高速化します。
- 代謝支持: 乳酸やクエン酸回路中間体を軸索へ供給し、ミエリン合成時の高エネルギー需要を支えます。
ミクログリア(微小膠細胞)
ミクログリアは脳内の免疫担当細胞で、安静時は枝分かれ状に静止しています。
- 監視・貪食: 異常タンパク質蓄積、死細胞、病原体を常時パトロールし、ファゴサイトーシス(細胞食作用)で除去します。
- シナプス刈込み: 発達期に過剰なシナプスを選択的に除去し、回路を最適化(経験依存的学習の基盤)。
- 炎症調節: サイトカイン(免疫シグナル分子)を放出し、感染時の免疫細胞動員を促進しますが、過剰活性化で慢性炎症を引き起こします。
末梢神経系での対応細胞
グリア細胞間のネットワーク機能
アストロサイトはCa²⁺波(カルシウムイオン濃度変動)を介して隣接細胞と通信し、局所血流増加や同期活動を調整します。これがfMRI(機能的磁気共鳴画像)で観察されるBOLDシグナルの基盤です。
役割比較表
| グリア細胞種類 | 主機能 | 分子機序 | 欠損疾患例 |
|---|---|---|---|
| アストロサイト | ホメオスタシス・シナプス制御 | グルタミン酸トランスポーター、グリア伝達物質 | てんかん、アルツハイマー病 |
| オリゴデンドロサイト | 高速信号伝達 | ミエリン基本タンパク(MBP)、PLP | 多発性硬化症 |
| ミクログリア | 免疫・刈込み | CX3CR1受容体、TREM2 | 神経変性疾患、自閉症 |
ポイント
- グリア細胞はニューロン依存ではなく独立シグナル伝達能を持ちます(グリア伝達)。
- 脳の90%を占めるにも関わらず、長年「支持細胞」と誤解されていましたが、現在は情報処理の積極的参加者と認識されています。
- グリア異常がうつ病、統合失調症、認知症の病態形成に寄与することが明らかになりつつあります。
神経細胞の静止電位は細胞の外部と比べ内部が一60mVである。動物細胞ではこの静止電位はどのように維持されているか。
静止電位の基本メカニズム
動物細胞の静止電位(細胞膜内外の電位差、約-60mV〜-70mV)は、主にイオン濃度勾配(細胞内外のK⁺、Na⁺濃度の差)と選択的イオン透過性(細胞膜がK⁺だけを通しやすい性質)によって維持されます。この電位差は、細胞が「静止」している状態で常に保たれ、活動電位(神経信号)の発生に必須です。
維持の2つの主要機構
1. Na⁺/K⁺-ATPase(ナトリウムカリウムポンプ)によるイオン濃度勾配の形成
細胞膜上のNa⁺/K⁺-ATPase(ATP分解酵素でイオンを能動輸送するタンパク質)は、以下の反応でイオン分布を維持します
- 3Na⁺を細胞外へ排出、2K⁺を細胞内へ取り込み(3:2の比率)。
- これにATP1分子を消費し、一次能動輸送として濃度勾配を作ります。
結果: - 細胞内:K⁺高濃度(約140mM)、Na⁺低濃度(約10mM)
- 細胞外:K⁺低濃度(約4mM)、Na⁺高濃度(約140mM)
この勾配が「イオン電池」の役割を果たします。
2. リークチャネル(漏洩チャネル)による静止電位の決定
細胞膜には常に開いているK⁺リークチャネル(カリウム漏洩チャネル)が多数存在します
- K⁺が濃度勾配に従って細胞外へ漏れ出す(拡散)
- K⁺(正電荷)が外へ出るため、細胞内が負に帯電(膜内側負電位)
- Na⁺リークチャネルも存在しますが、K⁺チャネルが支配的で、膜の選択的K⁺透過性により静止電位はK⁺平衡電位(EK ≈ -90mV)に近づきます(ゴールドマン方程式で計算)
- Na⁺/K⁺ポンプが漏れ出したイオンを補正し、平衡を保ちます
ゴールドマン・ホッジキン・カッツ方程式による定量的説明
静止電位(Vm)は以下の式で表されます:
- P:透過係数(PK >> PNa)
- [ ]o:細胞外濃度、[ ]i:細胞内濃度
これによりVm ≈ -60mV〜-70mVが導出されます。
動的平衡の維持プロセス
- K⁺リーク → 細胞内負電位形成
- 電気的力(負電位)がK⁺流出を抑制(ネルンスト平衡に近づく)
- Na⁺微小流入 → 電位が徐々に脱分極(正方向へ)
- Na⁺/K⁺ポンプがNa⁺排出・K⁺補充 → 勾配・電位を回復
このポンプとリークのバランスが静止電位を安定維持します
比較表:維持機構の要素
| 機構 | 役割 | 分子 | 効果 |
|---|---|---|---|
| Na⁺/K⁺ポンプ | イオン勾配形成 | ATPase酵素 | 一次能動輸送(ATP消費) |
| K⁺リークチャネル | 電位決定 | 漏洩型K⁺チャネル | K⁺拡散による分極 |
| 膜脂質二重層 | 選択的透過性 | リン脂質 | K⁺優位透過 |
ポイント
- 支配要因:K⁺リーク(70-90%寄与)、ポンプ(勾配維持)
- エネルギー消費:ポンプが細胞ATPの20-40%を消費
- 異常時:ポンプ阻害(ウアバイン)で電位消失、活動電位不能
- 意義:静止電位が活動電位の「基準線」となり、閾値(約-55mV)との差で興奮を検知
活動電位の三つの段階の名称を述べよ。それぞれの背景にある関連する分子基盤とイオンについて記述せよ。なぜ電位依存性チャネルという名称が活動電位の発生にかかわる Na+チャネルに使われているのか。
活動電位の3段階
活動電位(action potential、神経信号)は以下の3段階で進行します:
- 脱分極(Depolarization)
- 再分極(Repolarization)
- 後分極(After-hyperpolarization)
各段階の分子基盤とイオン動態
1. 脱分極(静止電位 -60mV → +40mV)
現象:膜電位が急速に正方向へ変化(脱分極)
分子基盤:
- 刺激により膜電位が閾値(約-55mV)を超えると、電位依存性Na⁺チャネル(voltage-gated Na⁺ channel)が開口します。
- チャネルの電圧センサー(S4セグメント:正電荷のアミノ酸配列)が電位変化を感知し、コンフォメーション変化(構造変化)を起こして開きます。
イオン動態:
- Na⁺が濃度勾配(外高内低)と電気的勾配(内負)を経て大量流入
- Na⁺平衡電位(ENa ≈ +60mV)に向かって電位が急上昇
2. 再分極段階(+40mV → -70mV)
現象:膜電位が負方向へ急速に戻る。
分子基盤:
- Na⁺チャネルの不活性化ゲート(ball-and-chain機構)が閉じ、Na⁺流入が停止
- 同時に電位依存性K⁺チャネル(voltage-gated K⁺ channel)が開口(活性化ゲート開く)
イオン動態:
- K⁺が濃度勾配(内高外低)に従って細胞外へ流出
- K⁺平衡電位(EK ≈ -90mV)に向かって電位が低下
3. 後分極段階(-70mV → -80mV)
現象:静止電位よりわずかに負になる過分極
分子基盤:
- K⁺チャネルの不活性化機構が遅延し、開口状態が持続
- Na⁺/K⁺-ATPase(ナトリウムカリウムポンプ)が徐々にイオン勾配を回復
イオン動態:
- K⁺流出が継続し、過剰分極
- 最終的にポンプ活動で静止電位(-60mV)へ回復
電位依存性チャネルの命名理由
「電位依存性(voltage-gated)」という名称は、チャネルの開閉が膜電位の変化に依存するためです。
分子構造の詳細:
- 各チャネルに電圧センサー領域(S4セグメント:アルギニンなどの正電荷残基が7番目のヘリックスに配列)があります。
- 脱分極(負→正)でS4が膜内で移動(外向き)、これが**チャネルポア(通路)**の開口を誘導します。
- 再分極でS4が内向きに戻り、チャネルが閉じる。
神経生理学的意義:
- この電位依存性により、静止電位では閉じ、閾値を超えると一斉開口(オール・オア・ナッシング則)。
- 活動電位の急峻な立ち上がり・立ち下がりと伝播性を保証します。
比較表:各段階のチャネル・イオン
| 段階 | 主チャネル | イオン | 電位変化 | 分子機構 |
|---|---|---|---|---|
| 脱分極 | 電位依存性Na⁺ | Na⁺流入 | -60mV→+40mV | 電圧センサー活性化 |
| 再分極 | 電位依存性K⁺ | K⁺流出 | +40mV→-70mV | 活性化ゲート開口 |
| 後分極 | K⁺(持続)+ポンプ | K⁺流出 | -70mV→-80mV | 遅延不活性化 |
ポイント
- 閾値依存:-55mVでNa⁺チャネルが「スイッチオン」(正帰還)。
- 不活性化機構:Na⁺チャネルの自動停止で持続興奮を防止。
- K⁺遅延:後分極で不応期を作り、連続発火を制御。
- 伝播の鍵:隣接膜を閾値以上に脱分極させる連鎖反応。
K+チャネルの結晶構造から、電位感受性ドメインがこのタンパク質の他の部分とどのように相互作用して開閉していると考えられているか、説明せよ。この構造ー機能相関は他の電位依存性イオンチャネルではどのように適用できるか。
要点
Kチャネルでは、電位感受性ドメイン(VSD;S1〜S4)の動きが、S4–S5リンカーを介して孔(pore;S5〜S6)のゲート(主にS6)へ機械的に伝わり、開閉が起きると考えられています。
この「VSD+孔」というモジュール構造と連結部でのカップリングという考え方は、他の電位依存性イオンチャネル(Na、Ca、HCNなど)にも基本的に適用できます。
結晶構造が示す部品配置
Kv(電位依存性K)チャネルは、各サブユニットがS1〜S6を持ち、S1〜S4がVSD、S5〜S6が中央の孔(pore)を作る、という設計になっています。
Kv1.2のX線結晶構造は、VSDが孔ドメインの周囲に配置され、電位センサーの動きが孔の開閉へ連結されうる“構造上の足場”があることを示しました。
またKvAPの構造でも、S1〜S4が孔の周りに配置され、S4を含む電位センサーの特徴的な配置(paddle様構造など)が議論されています。
VSD→ゲートの力の伝達(S4–S5リンカー)
結晶構造と機能解析を合わせた主流のモデルでは、膜電位変化でVSD(特にS4)が配置を変え、その変化がS4–S5リンカーへ伝わります。
S4–S5リンカーは、電位センサーと孔ドメインを“物理的につなぐ部品”で、内側ゲート(S6が作る束=bundle crossing)近傍に位置し、ゲート開閉に影響できる配置にあります。
PLOS Biologyの解析では、S4–S5リンカーが4本集まってカフ(cuff)のようにS6側を囲み、電位センサーの動きをゲートへ変換するテコ(lever)的役割を担う、という説明がまとめられています。
さらに、S4–S5リンカーがS6の束(bundle crossing)と同一サブユニット内・隣接サブユニット間の両方で相互作用しうることが、S4–S5リンカーの運動測定などから議論されています。
相互作用の候補は「1か所」ではない
VSDと孔の結合面はS4–S5リンカー周辺が代表的ですが、VSD(例:S1)と孔側の別の接点がありうることも示唆されています。
このように「複数の界面で、力の伝達や安定化が起きる」という見方を取ると、変異で開きやすさが変わる部位が複数ある理由を説明しやすくなります。
他の電位依存性チャネルへの適用
電位依存性チャネル一般に、S4の正電荷を含むVSDが膜電位を感知し、その動きが孔の開閉へ伝わる、という基本設計は共通モチーフとして扱われます。
NaチャネルやCaチャネルは1本のポリペプチドに4つの繰り返しドメイン(疑似4量体)としてVSD+孔のユニットを持つため、「VSDの動きが孔へ伝わる」という考え方自体はKvの枠組みで整理できます。
一方で、HCNなど一部チャネルでは電位センサーの動きとゲート開閉の向きやカップリング様式が異なる(同じ部品でも出力が違う)ため、Kvで確立したS4–S5リンカー中心の見方を出発点にしつつ、チャネルごとの例外(ドメイン配置やリンカー構造の違い)を検討する、という適用の仕方になります。
必要なら、ここでいう「開閉」を(A)S6束での内側ゲート(activation gate)とみなすのか、(B)選択性フィルター側の“別のゲート”も含めて述べるのか、どちらの教科書文脈かに合わせて書き分けます。
なぜ活動電位の強度が軸索の末端へと伝わっていく際に下がらないのか説明せよ。
受動的な電位変化は本来「減衰する」
膜電位変化(電流)が受動的に広がるだけなら、距離が離れるほど小さくなっていきます(いわば漏れながら伝わるため)ので、長い軸索の末端まで同じ強さで届きません。
減衰しない理由:各地点で「再生」される
活動電位は、ある場所で起きた脱分極が隣の膜を脱分極させ、そこで電位依存性Na⁺チャネルが開いて新たなNa⁺流入が起こり、同じサイズの活動電位が作り直される(再生される)ことで前へ進みます。
このため、伝わっているのは「同じ電位変化が弱まりながら流れていく」のではなく、「一定の大きさの活動電位が、場所を変えながら次々に発生している」という状態になります。
この性質は「全か無か(閾値を超えると一定の大きさで発生する)」という活動電位の特徴とも整合します。
逆流しないことも関係する(補足)
直前に活動電位が起きた領域は不応期(主にNa⁺チャネルの不活性化による)に入り、すぐには次の活動電位を起こせないため、後ろ向きに同じ強さで伝わることが抑えられます。
この「前方だけが発火できる」条件が保たれることで、再生の波が途切れにくく、末端まで安定して伝導します。
まとめ
活動電位が減衰しないのは、軸索の各区間で電位依存性Na⁺チャネルが開いて活動電位が繰り返し再生され、作り直されながら伝わる仕組みだからです。
神経シグナルが細胞体に向かって逆行することを、何が防いでいるのか。
活動電位が細胞体方向へ「逆行」しにくい最大の理由は、直前に興奮した膜領域が一時的に再び発火できない不応期に入り、後ろ向きの伝導が起きない(起きても広がらない)ためです。
加えて、神経回路としては化学シナプスが基本的に「シナプス前→シナプス後」の向きに作られているため、ニューロン間の情報の流れも一方向になりやすいです。
活動電位での防波堤
- Na⁺チャネル不活性化:活動電位の直後、電位依存性Na⁺チャネルは不活性化状態になり、同じ場所ではすぐに再びNa⁺流入を起こして発火できません(絶対不応期)。
- K⁺コンダクタンス増加:再分極〜後過分極の間はK⁺が出やすい状態が続き、膜電位が発火しにくい側へ引っ張られるため、逆向きの刺激が入りにくくなります(相対不応期)。
- この結果、「前方(まだ興奮していない領域)」は発火できる一方、「後方(さっき興奮した領域)」は発火できず、活動電位は前へ進む形になります。
シナプスでの一方向性(補足)
化学シナプスでは、神経伝達物質を放出する装置がシナプス前終末に、受容体が主にシナプス後膜側に配置されるため、情報は通常「前→後」に伝わります。
そのため、たとえ単一ニューロン内で局所的に逆行性の電気現象が起き得る状況があっても、回路全体としては細胞体へ逆流して同じ経路を戻る形にはなりにくいです。
「逆行」が(1)同一軸索内の逆向き伝導、(2)シナプスをまたいだ逆向き伝達、なのかで区別して説明することが必要です。
ミエリン形成によって軸索に沿った活動電位の伝播速度が上がる。ミエリン形成とは何か。ミエリン形成によって、軸索に沿って存在するランビエ絞輪における電位依存性 Na+チャネルと Na+/K+ポンプが集中化されている。ランビエ絞輪間の間隔を10倍に増すと、活動電位の伝播にどのような影響が出ると予想されるか。
ミエリン形成とは何か
ミエリン形成とは、グリア細胞が軸索の周りに自分の膜を何重にも巻き付けて「ミエリン鞘(myelin sheath)」という絶縁層を作ることです。
この巻き付けられた区間(internode)と巻き付けが途切れる短いすき間(ランビエ絞輪;node of Ranvier)が、軸索に沿って繰り返し並びます。
末梢神経ではシュワン細胞、 中枢神経ではオリゴデンドロサイトがミエリンを作る、という役割分担が一般的です。
なぜ伝播速度が上がるか
ミエリン鞘があると、活動電位は軸索全体を連続的に再生しながら進むのではなく、ランビエ絞輪から次の絞輪へ跳ぶように進む(跳躍伝導;saltatory conduction)形になります。
この方式だと、活動電位として「作り直し(再生)」が必要なのは主に絞輪部分なので、同じ距離を進むのに必要な再生ポイントが減り、結果として速く見えます。
絞輪にNa⁺チャネルとNa⁺/K⁺ポンプが集まる意味
ランビエ絞輪では電位依存性Na⁺チャネルが高密度に集積しており、ここで強いNa⁺流入を起こして活動電位を再生することが、速く確実な伝導に必要だと考えられています。
中枢神経の有髄軸索では、Na⁺チャネルやK⁺チャネルの「局在化・クラスター化(集まって配置されること)」が、適切な信号伝達に重要だという整理もされています。
また、絞輪ではNa⁺/K⁺-ATPase(Na⁺/K⁺ポンプ)も機能的に重要で、活動電位で流入したNa⁺や流出したK⁺によって崩れたイオン勾配を回復して、繰り返し発火できる状態を支えます。
絞輪間隔を10倍にするとどうなるか(予想)
絞輪間隔(internode長)が長くなると、次の絞輪を閾値まで脱分極させるための電流がより遠くまで届く必要があり、その途中で電位変化が減衰しやすくなるため、伝導の安全率(確実性)が下がる方向に働くと予想されます。
したがって、10倍の増加は「速くなる」よりも、むしろ 伝導が遅くなる、あるいは条件によっては次の絞輪が十分に脱分極できず 伝導が途切れる(伝導ブロック)リスクが上がる、という影響が出る可能性があります。
一方で、モデルや条件によっては「internode長を増やすと伝導速度が上がる」ケースも報告されており、絞輪間隔には長いほど良いではなく、軸索の太さ・膜特性・外液抵抗などと釣り合った最適域がある、という発想で理解すると整理しやすいです。
まとめ
- ミエリン形成は、グリアが軸索を絶縁して、絞輪で活動電位を再生する「跳躍伝導」を可能にします。
- 絞輪へのNa⁺チャネル集中は再生のため、Na⁺/K⁺ポンプはイオン勾配回復のために重要です。
- 絞輪間隔を10倍にすると、一般には安全率が下がり、遅延や伝導ブロックのリスク増加が予想されます(ただし条件依存で最適域の考え方が必要です)。
アセチルコリンはシナプスで放出される一般的な神経伝達物質である。神経筋シナプスにおいてアセチルコリンエステラーゼの活性が低下した筋肉活動はどのような結果になるか、予測せよ。
筋肉活動の予測
神経筋シナプスでアセチルコリンエステラーゼ(AChE)の活性が低下すると、放出されたアセチルコリン(ACh)が分解されにくくなり、シナプス間隙にAChが残りやすくなります。
その結果、筋細胞側の受容体が刺激され続け、筋収縮の「オン」が切れにくくなって、筋の収縮が長引いたり、異常な筋の緊張・けいれん様の状態が起こりやすくなると予想されます。
強く長くAChE活性が落ちる状況では、最終的には筋がうまく力を出せなくなる(麻痺・脱力)方向の影響も起こりえます。
そうなる理由
AChEは、シナプス間隙に出たAChを速やかに分解して、神経から筋への信号を「止める」役割を担います。
この止める仕組みが弱まると、次の信号が来ていないのに受容体が刺激され続けるため、筋側では本来より長い時間、興奮(脱分極)や収縮が続く方向にずれます。
さらに刺激が続くと、筋側は正常な「収縮→弛緩→次の収縮」というリズムを作りにくくなり、結果として運動がぎこちなくなったり、力が入らなくなったりする、と考えると整理しやすいです。
まとめ
AChE活性低下は「AChが分解されずに残る」ことを通じて、神経筋シナプスの信号終結を遅らせ、筋活動を過剰・持続化させ、状況によっては麻痺(脱力)にもつながると予測できます。
刺激された細胞で活動電位がシナプスに到達すると、シナプス小胞は速やかにシナプス前細胞の細胞膜に融合する。これは1ミリ秒以内に起こる。このように驚くべき速さでこの現象が起こるのはどのような機構によるか。
何が「1ミリ秒以内」を可能にするか
この現象が速い本質は、活動電位が来てから融合の準備を始めるのではなく、融合直前の状態まで事前にセットしておき、最後の引き金だけをCa²⁺で引く設計になっている点です。
実際、活動電位→電位依存性Ca²⁺チャネルからのCa²⁺流入→Ca²⁺センサーの作動→小胞融合、という一連が1ms未満で起きうることが議論されています。
事前準備:ドッキングとプライミング
放出に使われる小胞の一部は、シナプス前終末の「活性部位(active zone)」で、膜に接近した放出準備完了(release-ready)の集団として保持されます。
この準備段階では、SNARE複合体(小胞側と細胞膜側の膜融合装置)が組み上がる方向へ進み、Munc13などがその“プライミング(放出可能化)”を進める仕組みが重要だと整理されています。
つまり、活動電位が到達した時点で「小胞はすでに現場に並び、分子機械も半分以上組み立て済み」なので、残りの工程が非常に短時間で済みます。
引き金:Ca²⁺ナノドメイン(距離が近い)
活動電位が到達すると、シナプス前膜の電位依存性Ca²⁺チャネルが開いて局所的にCa²⁺が流入し、細胞内Ca²⁺濃度が短時間だけ急上昇します。
速いシナプスでは、Ca²⁺チャネルとCa²⁺センサー(後述)がナノメートルスケールで近接し、「Ca²⁺ナノドメイン」による強い局所シグナルで放出が引き起こされる、と説明されています。
この“距離の近さ”により、Ca²⁺が細胞内に広がって薄まる前に、必要な分子へ即座に届くため、タイムロスが最小化されます。
融合:SNARE+シナプトタグミン(最終スイッチ)
膜融合の力そのものは、SNAREタンパク質が「ジッパーのように」結合を進めて膜同士を引き寄せることで生まれる、というモデルが中心です。
ただしSNAREだけだと勝手に融合しやすく制御が弱いため、神経終末ではシナプトタグミン(synaptotagmin)がCa²⁺センサーとして働き、Ca²⁺流入を合図にSNARE機構へ介入して融合を一気に進める役を担う、と整理されています。
さらに複雑な制御として、シナプトタグミンやコンプレキシン(complexin)が融合を抑えた待機状態(クランプ)を作り、Ca²⁺結合をきっかけにその抑制が外れて同期的な高速放出が起こる、という見方も提示されています。
まとめ
1 ms以内の速い小胞融合は、「小胞とSNARE装置を活性部位で事前に準備」し、「Ca²⁺チャネルとセンサーを極近距離に配置」して、「Ca²⁺結合でクランプ解除→SNAREが最終ジッパリング」という流れで最終工程だけを一瞬で走らせることで実現します。
脳においては特に、神経細胞は複数の興奮性と抑制性のシグナルを受け取る。神経細胞のこのシグナルを受け取る突起の名前は何か。神経細胞はこのシグナルをどのように統合し、活動電位を生じさせるか否かを決定しているか。
シグナルを受け取る突起の名前
神経細胞が他の細胞からの入力(シナプス入力)を主に受け取る突起は、樹状突起です。
興奮性シナプスが樹状突起に投射する場合、樹状突起の表面に「樹状突起棘(dendritic spine)」というトゲ状の小突起が作られることがあります。
神経細胞には軸索と樹状突起という2種類の突起があり、一般に軸索側が出力、樹状突起側が入力を担います。
興奮性・抑制性シグナルの統合
脳のシナプスには興奮性シナプスと抑制性シナプスがあり、興奮性シナプスは脱分極方向の変化(興奮性シナプス電位:EPSP)を、抑制性シナプスは発火しにくくする方向の変化(抑制性シナプス電位:IPSP)を起こします。
神経細胞は、樹状突起や細胞体に生じた多数のEPSPとIPSPを「足し算」して統合し、最終的に活動電位を出すかどうかを決めます。
この加算には、同時刻に別々の場所から来た入力を足す「空間的加算」と、短時間に繰り返し来た入力を足す「時間的加算」があります。
活動電位を出すかの決定
統合の結果、膜電位がしきい値に達すると活動電位が発生し、達しなければ発生しません。
多くのニューロンでは、この最終判断が「軸索起始部(軸索の出発点付近)」で行われる、と理解すると整理しやすいです。
抑制性入力は、膜電位をより負に寄せてしきい値から遠ざけるだけでなく、膜の電気的性質を変えて興奮性入力の効き目を弱める(シャント抑制)ことで、発火を起こりにくくする場合もあります。
まとめ
入力を受け取る主役は樹状突起(必要に応じて樹状突起棘)で、興奮性・抑制性のシナプス電位を加算した結果がしきい値を超えるかどうかで、活動電位を出すかが決まります。
シナプス小胞の再利用において、ダイナミンはどんな役割をしているか。それを支持する証拠は何か。
ダイナミンの役割
ダイナミン(dynamin)はGTPを使って働く酵素(メカノケミカルGTPase)で、シナプス小胞が膜融合で放出した後、その膜を「取り戻して新しい小胞に戻す」過程(エンドサイトーシス)で重要な役割を担います。
特に、細胞膜が内側へへこんでできたくびれ(首)の部分で、ダイナミンが集まって締め付け、芽のようにできた小胞を膜から切り離す(膜分裂/scission)工程に関与すると考えられています。
言い換えると、ダイナミンは「小胞を作りかけの状態(膜にぶら下がった状態)」を「独立した小胞」に仕上げる最後の切り離し係です。
なぜ再利用に必須か
神経終末では、神経伝達物質を入れた小胞が放出(エキソサイトーシス)されるたびに小胞膜成分が細胞膜へ足されるので、同じ分だけ回収して小胞を作り直さないと、放出を繰り返せません。
ダイナミンが働かないと、この回収(補償性エンドサイトーシス)が止まり、結果として使える小胞の補充が追いつかず、刺激を繰り返したときに神経伝達が続かなくなります。
それを支持する証拠
温度感受性のショウジョウバエ変異体 shibire(ダイナミンをコードする遺伝子)が高温条件で機能不全になると、シナプス小胞リサイクリングが障害されることが示されています。
このとき神経終末を電子顕微鏡で観察すると、シナプス小胞が枯渇し、同時にクラスリン被覆ピット(coated pits)が多数たまる(=くびれたまま切り離されない構造が増える)ことが報告されています。
また、ダイナミン阻害剤(例:dynasore)を用いると補償性エンドサイトーシスが強く阻害され、刺激を続けるほど放出イベントが減っていく(小胞が補充されない)ことが示されています。
さらに、ダイナミンとシナプトフィジンの相互作用を乱す操作(ペプチド注入)により高頻度刺激で神経伝達が落ち、電子顕微鏡で小胞回収がうまくいかない所見(小胞の減少など)が見える、という報告もあります。
まとめ
ダイナミンは、回収中の膜を最終的にちぎって独立した小胞として再生させる段階に深く関わり、その阻害や変異で「回収途中のピットがたまる」「小胞が枯渇して伝達が続かない」という現象が観察されることが、役割の強い証拠になります。
桿体細胞や錐体細胞が光によって刺激されたとき、一連の現象がカスケードとして実行され、それは最終的に視覚情報として脳へ伝達される。ロドプシンによる光吸収から桿体と錐体の細胞膜の過分極に至るこのカスケードの分子的な過程を概説せよ。
ロドプシンの光吸収(開始)
桿体・錐体の視物質(桿体ではロドプシン)は、光を吸収すると補欠分子(レチナール)が 11-cis 形から all-trans 形へ異性化し、ロドプシン(オプシン)が活性化状態へ構造変化します。
この「光でGPCR(Gタンパク質共役受容体)が活性化される」という点が、視覚の分子カスケードの出発点です。
トランスデューシン活性化(増幅)
活性化ロドプシンはGタンパク質トランスデューシン(transducin)を次々に活性化し、GαサブユニットがGDP→GTP交換してシグナルを運びます。
ここで「1つの光刺激が複数の分子反応を連鎖的に進める」ため、信号が増幅されます。
PDE6がcGMPを減らす(セカンドメッセンジャー低下)
活性化トランスデューシンはホスホジエステラーゼ6(PDE6)を活性化し、細胞内のcGMPを加水分解して濃度を下げます。
PDE6活性化によるcGMP低下が、膜電位変化へ直結する中心のスイッチです。
CNGチャネル閉鎖→過分極(ゴール)
暗所ではcGMPが比較的高く、cGMP依存性陽イオンチャネル(CNGチャネル)が開いてNa⁺やCa²⁺が流入する「暗電流(dark current)」が流れています。
光でcGMPが減るとCNGチャネルが閉じ、陽イオンの流入が止まって暗電流が減少するため、細胞膜は過分極(より負になる)します。
この過分極により、シナプス終末からのグルタミン酸放出が減少し、その変化が網膜回路で処理されて最終的に脳へ伝えられる、という流れになります。
まとめ
光→(レチナール異性化で)ロドプシン活性化→トランスデューシン→PDE6→cGMP低下→CNGチャネル閉鎖→暗電流低下→過分極、が「ロドプシンから過分極まで」の分子カスケードです。
視覚におけるコントラスト(差異)の検出にはどのような細胞がかかわっているか。これらの細胞は、コントラストを検出するために光受容細胞の情報をどのように統合しているか。
コントラスト検出に関わる細胞
コントラスト(明暗の差、境界)を脳へ出力する中心的な細胞は、網膜の神経節細胞(retinal ganglion cell; RGC)です。
RGCへ信号を渡す中継として、双極細胞(bipolar cell)が光受容細胞(桿体・錐体)からの信号を受けてRGCへ伝えます。
さらに、コントラスト検出に重要な横方向の抑制(lateral inhibition)を作る細胞として、水平細胞(horizontal cell)とアマクリン細胞(amacrine cell)が関わります。
光受容細胞の情報の統合方法(中心・周辺と側方抑制)
コントラスト検出の基本設計は、多くの細胞が「中心(center)と周辺(surround)が拮抗する受容野(center-surround receptive field)」を持つことです。
この拮抗(中心が興奮なら周辺は抑制、またはその逆)があると、網膜回路は絶対的な明るさよりも明暗の境界に強く反応しやすくなります。
側方抑制による中心・周辺の受容野構造は、RGCが空間的・時間的コントラストに敏感になるための仕組みだと整理されています。
どこで「周辺抑制」が作られるか
周辺(surround)側の抑制成分は、網膜の複数の層で作られうるとされ、少なくとも水平細胞とアマクリン細胞が周辺成分に寄与することが示されています。
水平細胞は光受容細胞からの入力を横方向に統合し、光受容細胞や双極細胞への抑制性フィードバック/側方相互作用を通じてコントラスト増強(lateral inhibition)に関わると説明されています。
実際に、水平細胞の機能を欠くとRGCの中心・周辺受容野の形成が障害され、空間的なチューニング(周辺の拮抗)が弱くなることが報告されています。
アマクリン細胞もRGCの周辺成分に寄与しうることが示されており、水平細胞とアマクリン細胞が異なる特徴の周辺成分を作ってRGCの受容野を形作る、という見方があります。
まとめ
コントラスト検出は、双極細胞・水平細胞・アマクリン細胞が光受容細胞の信号を統合して「中心・周辺が拮抗する入力」を作り、それを神経節細胞がスパイクとして脳へ出力することで実現します。
カエルにおける Roger Sperry の実験では、神経細胞がどのように接続されていくかを説明するための共鳴仮説と化学親和仮説をどのように区別したか。
争点:2つの仮説の違い
共鳴仮説(Weiss らの立場)は、軸索の結合は最初から厳密に相手が決まっているというより、機能的な相互作用や活動(使われ方)に合わせて結線が調整される、という考え方です。
化学親和仮説(Sperry)は、軸索と標的細胞の側に分子レベルのタグがあり、その特異的な相互作用で正しい相手が選ばれて結合が決まる、という考え方です。
Sperryがした操作(眼球180°回転+視神経切断)
Sperry はカエルで、眼球を180度回転して移植し直した上で視神経を切断し、視神経が再生した後の見え方・行動がどうなるかを調べました。
この実験は「網膜—視蓋(tectum)投射が、回転後の見えに合わせてつなぎ替わるのか、それとも回転前の対応関係に従ってつながるのか」を直接テストする設計になっています。
観察結果:再生後も上下が逆の行動
視神経が再生した後も、カエルの行動は回転後の網膜像に合わせて修正されず、餌の位置に対して不適切な向きの定位・捕食行動が続く(例:上の餌に下へ反応するようなズレが残る)ことが示されました。
この結果は「再生した軸索が、眼球の回転に合わせて新しい対応へ学習・調整してつなぎ替わった」という説明と合いにくい所見です。
どう区別できたか(ロジック)
もし共鳴仮説(活動・機能に合わせた結線調整)が主要因なら、回転後の視覚入力と成功する行動の対応に合わせて結線が組み替わり、時間とともに行動のズレが小さくなることが期待されます。
しかし実際には、回転後も回転前に対応していた視蓋領域へ投射が再形成されたと解釈できる行動が持続したため、「標的選択に分子タグの特異性がある」という化学親和仮説の方が説明しやすい、という結論につながりました。
脳科学辞典でも、この一連の結果から Sperry は軸索と標的細胞の分子タグの特異的相互作用がトポグラフィックマップ形成に関与すると提唱した、とまとめられています。
まとめ
眼球回転という入力座標を入れ替える操作をしたのに、再生後の投射がそれに合わせて再配線されず、元の対応に従って結合したと解釈できる結果が出たことで、Sperry は共鳴仮説より化学親和仮説を支持しました。
問題文引用元:東京化学同人 分子細胞生物学 第6版


コメント