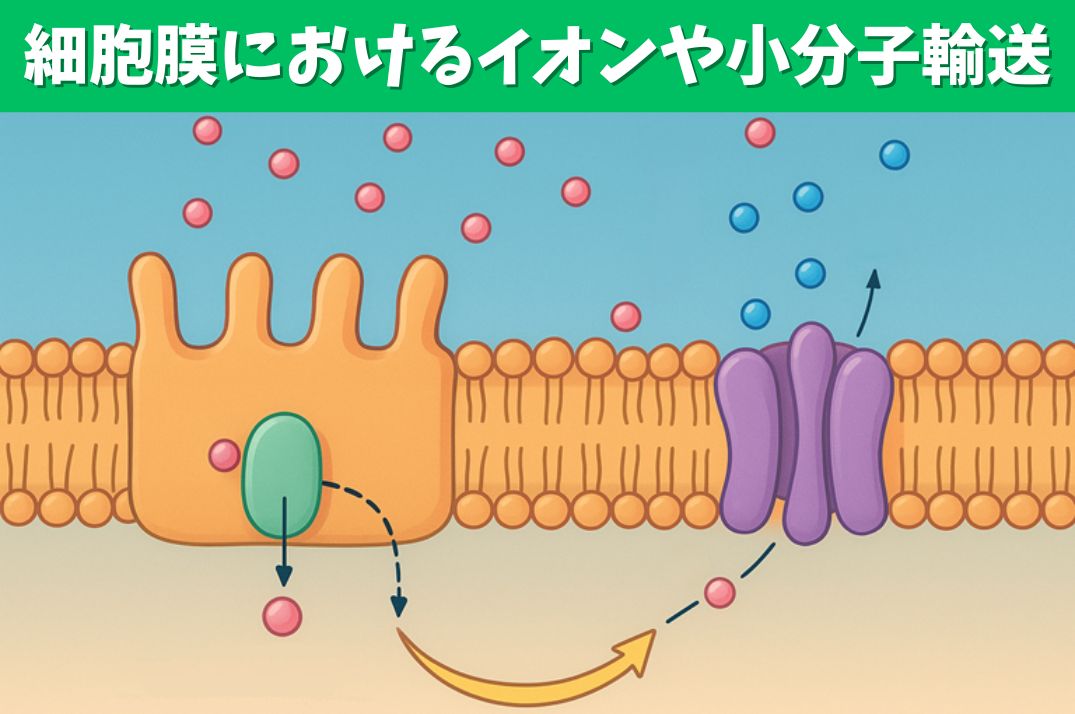
- 生体膜の基本構造はリン脂質二重層である。酢酸とエタノールはともに2個の炭素そして水素と酸素からなり、どちらも受動拡散で細胞内に入る。pH7のとき、どちらか一方がより膜を透過しやすい。それはどちらか、そしてなぜか。典型的な胃のpHであるpH1.0にしたらそれぞれの透過性はどうなるか。
- 2.単一輸送体とイオンチャネルは生体膜で促進拡散を行っている。どちらも促進拡散の例としてあげられるが,チャネルを通る速度のほうが単一輸送体より $ 10^4~10^5$倍大きい。どのような機構上の違いがこうした大きな輸送速度の違いを生むのか。
- 3種類の輸送体の名称をあげよ。そのうちのどれがグルコースあるいは炭酸水素イオンHCO3-を電気化学的配に逆らって輸送することができるか。グルコースではそうではないが、炭酸水素イオンの場合,輸送過程のΔGは二つの項を含む。その二つの項とは何で、第二の項はなぜグルコース輸送にはないのか。これらの共輸送体はしばしば二次的能動輸送の例としてあげられる。それはなぜか。
- 赤血球細胞膜にある GLUT1は単一輸送体の古典的例である。GLUT1がガラクトースやマンノースではなくグルコースにだけ特異的な輸送体であることを証明する実験計画をたてよ。グルコースは六炭糖でリボースは五炭糖である。リボースはグルコースより小さいのにGLUT1により輸送されにくい。どのようにこれを説明するか。
- イオンや分子を能動輸送するATP駆動ポンプには4種類ある。それらの名称をあげよ。それらのうちのどれがイオンだけを輸送し、どれがおもに小分子を輸送するのか。これらのATP駆動ポンプのうちの一つは、天然の基質ではなくがんの化学療法のための人工基質の輸送について調べているときに発見された。現在、研究者はこの特別なATP 駆動ポンプの天然基質はどのようなものだと考えているのか。
- ゲノム解読は続き、ゲノムの塩基配列がすべて明らかになった生物も増えている。こうした情報からマウスあるいはヒトに特定の輸送体またはポンプが何個存在するかを調べるにはどうしたらよいか。塩基配列から同定された輸送体またはポンプの多くは、その天然の基質や生理的役割がわからないという意味で“オーファン”タンパク質である。どうしてそうなるのか。こうした“オーファン”タンパク質の生理的役割を明らかにするにはどうしたらよいのか。
- 特定のチャネルやポンプや輸送体に対する阻害剤や活性化剤は製薬業界が産生する薬のかなり大きな部分を占めている。骨格筋の収縮は細胞質Ca2+濃度上昇によってひき起こされる。筋小胞体(SR)のP型Ca2+ ATPaseを選択的に阻害する薬物は筋収縮にどのような影響を与えると考えられるか。
- 植物とは違い、動物細胞の膜電位はおもに静止K⁺チャネルによって生じる。これらのチャネルはどのようにして静止膜電位形成に寄与するのか。なぜこれらのチャネルは開閉調節を受けないとみなされているのか。これらのチャネルはどのようにK⁺とNa⁺を区別しているのか。
- パッチクランプ法により個々のイオンチャネルの電気伝導度を測定することができる。K⁺チャネルと思われる遺伝子産物が実際にK⁺あるいは Na⁺チャネルであることを確認する上でパッチクランプ法はどのように使われるかを述べよ。
- 植物は液胞膜をはさんでのH⁺の電気化学的配を使って液胞に塩や糖をたくわえている。これにより液胞は高張になる。植物細胞はなぜこれによって破裂しないのか。動物細胞膜のN⁺/K⁺ATPase は張条件下での動物細胞の破裂を防いでいる。どのようにしてそれを行っているのか。
- 細菌の Na⁺/ロイシン輸送体の場合、結合イオンをK⁺ではなく Na⁺にしていることの重要な点は何か。小腸上皮細胞においてグルコースを等方輸送するしくみを説明せよ.この輸送に使われるイオンは何で、このイオンが細胞膜を通って流入することがエネルギー的に好都合となる理由を二つあげよ。
- ゼブラフィッシュなどいくつかの生物のゲノム塩基配列が解明され、多数の受容体や輸送体の存在が明らかになった。ゼブラフィッシュのgolden 遺伝子をポジショナルクローニングによって調べたところ、陽イオン交換タンパク質と思われる SLC24A5であることがわかった。このgoldlen遺伝子に突然変異が起こると、メラニンという黒色色素の量が大きく減るので、通常水平方向に走っている黒い縞が薄くなり、全体が金色に見えるようになる、魚類やヒトにあるメラニンを含む膜小胞は何とよばれるか。このgolden 遺伝子からつくられるタンパク質がゼブラフィッシュのどこで発現されどこに局在するのかを明らかにする実験計画を立案せよ。このゼブラフィッシュのgolden 遺伝子に突然変異が起こったときに、他の動物のオルソロガス遺伝子を発現させて、その表現型をもとに戻すことを何というか。
- 小腸上皮細胞での内腔側から血液側へのグルコース輸送は経細胞輸送の重要な例である。Na⁺/K⁺ATPase はどのようにこの過程の原動力となっているのか。この過程に密着結合が重要なのはなぜか。スポーツドリンクのような水分補給のための飲料は糖と塩を含んでいる。なぜその二つが水分補給に重要なのか。
生体膜の基本構造はリン脂質二重層である。酢酸とエタノールはともに2個の炭素そして水素と酸素からなり、どちらも受動拡散で細胞内に入る。pH7のとき、どちらか一方がより膜を透過しやすい。それはどちらか、そしてなぜか。典型的な胃のpHであるpH1.0にしたらそれぞれの透過性はどうなるか。
1. 酢酸とエタノールの透過性比較(pH7で)
- エタノールのほうが膜を透過しやすいです。
- 理由:
- エタノールは非イオン化(中性)分子であり、分子全体が比較的親脂性・疎水性であるため、脂質二重層を容易に通過できます。
- 酢酸は弱酸性の化合物で、pH7環境では主にイオン化された酢酸イオン(CH3COO⁻)の形態で存在します。
- イオン化した酢酸は親水性が強く、疎水性の膜内部を透過しにくいので透過性が低くなります。
2. pH1.0(胃の典型的pH)での透過性変化
- pHが非常に低い(酸性)環境では、酢酸は主に非イオン化の酢酸分子(CH3COOH)として存在します。
- 非イオン化の酢酸はエタノールのように疎水的性質が強くなるため、酢酸の膜透過性が高まる。
- エタノールはもともと非イオン化でpHによる影響はほとんど受けないため、透過性はほぼ変わりません。
まとめ
| 条件 | 膜透過性が高い物質 | 理由 |
|---|---|---|
| pH7 | エタノール | 酢酸はイオン化して親水性が高く、膜を通りにくい |
| pH1.0 | 酢酸とエタノール | 酢酸は非イオン化され疎水性になり透過性が向上。エタノールは変わらず |
つまり、pHによって酢酸のイオン化状態が変わり、その疎水性が上がることで膜透過性が増し、pH7ではエタノールが透過しやすいのに対し、pH1.0では酢酸も高い透過性を示すようになります。
2.単一輸送体とイオンチャネルは生体膜で促進拡散を行っている。どちらも促進拡散の例としてあげられるが,チャネルを通る速度のほうが単一輸送体より $ 10^4~10^5$倍大きい。どのような機構上の違いがこうした大きな輸送速度の違いを生むのか。
1. 基本的な違い
- 単一輸送体(キャリアタンパク質)は、基質(イオンや小分子)を結合し、膜を横断するために構造変化(コンフォメーションチェンジ)を繰り返す必要があります。
- イオンチャネルは膜を貫通する開いた孔(通路)を形成し、イオンや水分子が自由に高速で流れるため、輸送速度が極めて速い。
2. 輸送速度の違いの機構的原因
- 輸送体の速度制限
単一輸送体は、基質結合→運搬→解離という一連の段階的プロセスを経るため、それぞれのステップごとに時間がかかります。1つの輸送体が1秒間に輸送できる分子数は限られる。ゆえに移動速度は比較的遅い。 - チャネルの高速輸送
イオンチャネルはイオンに対して水のような通路を提供し、基質は自由に電気化学勾配に従って流れます。- チャネルは多くのイオンを同時に「パイプ」状の通路で通すことが可能で、輸送速度は拡散速度に近い(1秒間に百万〜千万個のイオンが流れる)。
- 基質結合や構造変化の必要がないため、時間的ロスがほぼない。
3. まとめ
| 特徴 | 単一輸送体 | イオンチャネル |
|---|---|---|
| 輸送原理 | コンフォメーションチェンジを伴う結合と運搬 | 開いた孔を通る自由拡散 |
| 輸送速度 | 1秒間に数千分子程度 | 1秒間に百万〜千万分子程度 |
| 特徴 | 逐次輸送なので速度が制限される | 複数のイオンが同時通過可能で高速 |
| 基質の選択性と制御性 | 高い(基質特異性が強い) | 高い選択性を持つが、速い流れを実現 |
このように、構造変化の有無と通過通路の性質の違いが輸送速度の桁違いの差を生んでいるのです。
3種類の輸送体の名称をあげよ。そのうちのどれがグルコースあるいは炭酸水素イオンHCO3-を電気化学的配に逆らって輸送することができるか。グルコースではそうではないが、炭酸水素イオンの場合,輸送過程のΔGは二つの項を含む。その二つの項とは何で、第二の項はなぜグルコース輸送にはないのか。これらの共輸送体はしばしば二次的能動輸送の例としてあげられる。それはなぜか。
1. 3種類の輸送体の名称
- チャネル(Channel)
物質やイオンが孔(チャネル)を通って受動的に拡散する。 - 単一輸送体(Carrier/Uniporter)
特定の分子を膜の一方から他方へ受動的に輸送する。例:グルコース輸送体GLUT。 - 共輸送体(Cotransporter / Symporter or Antiporter)
2種類以上の物質を同時に輸送する。- シンポーター(共輸送):同じ方向に輸送
- アンチポーター(交換輸送):逆方向に輸送
2. 電気化学的勾配に逆らって輸送できるのは?
- グルコースの場合
- グルコースは通常チャネルや単一輸送体(例:GLUT)で、電気化学的勾配なしに拡散に従うので「逆らって輸送する」ことはない。
- 炭酸水素イオン(HCO3⁻)の場合
- HCO3⁻は電荷を持つため電気化学的勾配が輸送に影響する。
- それを逆らう輸送は共輸送体(多くはアンチポーターやシンポーター)で可能であり、エネルギー源として他のイオン勾配(通常Na⁺やH⁺)を利用する。
3. 炭酸水素イオン輸送のΔGが含む2つの項とは?
炭酸水素イオンの輸送に伴う自由エネルギー変化ΔGは以下の2つの項で表されます。
- 化学的勾配成分(濃度差による)
$$
\Delta G_{\mathrm{chem}} = RT \ln \left( \frac{[\mathrm{HCO_3}^-]{\mathrm{in}}}{[\mathrm{HCO_3}^-]{\mathrm{out}}} \right)
$$
- 電気的勾配成分(膜電位差による)
$$
\Delta G_{\mathrm{elec}} = zF \Delta \Psi
$$
($z$:イオン価数, $F$:ファラデー定数, $\Delta \Psi$:膜電位差)
4. 第二の項(電気的勾配成分)がグルコース輸送ではない理由
- グルコースは電気的に中性分子であり、膜電位差の影響を受けないため、電気的勾配成分(第二の項)がありません。
- したがって、グルコースの受動的輸送は濃度勾配のみに従います。
5. なぜ共輸送体が二次的能動輸送の例なのか
- 二次的能動輸送(Secondary active transport)はATPを直接使わずに、他のイオン(例:Na⁺やH⁺)の電気化学的勾配を利用して別の物質を逆らって輸送する仕組みをいいます。
- 共輸送体はこの方式で、例えばNa⁺勾配のエネルギーを使ってHCO3⁻を細胞内に引き込むなど、エネルギー的に不利な方向への輸送を実現します。
- これに対し、単一輸送体はATPも使わず、物質を自然な勾配に沿って受動拡散させるだけです。
まとめ
| 輸送体の種類 | 逆勾配輸送可能か | ΔGの項 | 備考 |
|---|---|---|---|
| チャネル | できない | 濃度勾配のみ | 速い拡散路 |
| 単一輸送体(GLUT) | できない | 濃度勾配のみ | グルコースの受動輸送用 |
| 共輸送体(例:Na⁺/HCO3⁻) | できる | 濃度勾配+電気勾配 | 二次的能動輸送として機能 |
このように、共輸送体は膜電位差を含むイオン勾配のエネルギーを利用し、電気化学的勾配に逆らった輸送を可能にしているため、重要な能動輸送機構となっています。
赤血球細胞膜にある GLUT1は単一輸送体の古典的例である。GLUT1がガラクトースやマンノースではなくグルコースにだけ特異的な輸送体であることを証明する実験計画をたてよ。グルコースは六炭糖でリボースは五炭糖である。リボースはグルコースより小さいのにGLUT1により輸送されにくい。どのようにこれを説明するか。
1. GLUT1のグルコース特異性を証明する実験計画
実験概要
赤血球細胞膜のGLUT1による糖輸送の特異性を検証するため、放射性や蛍光標識したグルコース、ガラクトース、マンノースの取り込みを比較的に測定します。
手順
- 細胞/膜の準備
純化したヒト赤血球、またはGLUT1を発現する細胞株を用いる。 - 標識糖の準備
- 放射性同位元素で標識された[14C]-グルコース、[14C]-ガラクトース、[14C]-マンノースを用意
- それぞれの標識濃度を揃える - 糖輸送の測定
- 同一条件で細胞に標識糖を添加し、一定時間(例:数分)後に細胞内への取り込み量を測定(放射能カウント)
- 無標識のグルコースを多量に添加した競合実験も行い、特異的輸送を確認 - 比較と解析
標識グルコースの取り込み量が標識ガラクトースやマンノースより有意に高ければ、GLUT1のグルコース特異性を示す証拠となる。
補足
- 競合阻害実験で、グルコースは輸送を抑制するが、ガラクトースやマンノースは抑制しないことも確認する。
- 補足実験として、GLUT1をノックアウトまたは阻害した条件下で糖輸送が減るかも確認する。
2. リボースがグルコースより輸送されにくい理由の説明
- グルコースは六炭糖(ヘキソース)、リボースは五炭糖(ペントース)であり分子構造が大きく異なる。
- GLUT1は特に六炭糖の形に合った立体的な結合部位(基質特異性)を持っていて、リボースの形(環のサイズやヒドロキシ基の配置)が適合しないため、輸送効率が非常に低い。
- 小さいリボースでも、形状や化学構造の違いにより受容部位との結合親和性が低く、結果として通過しにくい。
まとめ
- GLUT1のグルコース特異性は標識糖を用いた取り込み実験や競合阻害実験で証明できる。
- 輸送の選択性は糖の分子構造・立体構造によって決まり、リボースはその特異的基質に合わないため、たとえ小さくても輸送されにくい。
この実験設計によりGLUT1の基質選択性の理解と機序解析に繋がります。
イオンや分子を能動輸送するATP駆動ポンプには4種類ある。それらの名称をあげよ。それらのうちのどれがイオンだけを輸送し、どれがおもに小分子を輸送するのか。これらのATP駆動ポンプのうちの一つは、天然の基質ではなくがんの化学療法のための人工基質の輸送について調べているときに発見された。現在、研究者はこの特別なATP 駆動ポンプの天然基質はどのようなものだと考えているのか。
1. ATP駆動ポンプの4種類の名称
- P型ATPアーゼ
- F型ATP合成酵素
- V型ATPアーゼ
- ABC輸送体(ATP-binding cassette transporter)
2. イオンを輸送するのはどれか
- P型ATPアーゼとV型ATPアーゼ
- P型はNa⁺/K⁺ポンプ、Ca²⁺ポンプなどのイオン輸送ポンプ
- V型は主にリソソームや細胞内小器官の膜に存在し、H⁺イオンを輸送
- F型ATP合成酵素は主にプロトンを膜を通して輸送し、ATP合成を担う。厳密には輸送というより逆反応でATP作製。
3. 小分子を輸送するのはどれか
- ABC輸送体
- 多様な基質(薬剤、脂質、ペプチド、小分子など)を輸送
- 複数の多重膜貫通サブユニットとATP結合タンパク質を持つ。
4. がん化学療法の人工基質の輸送調査から発見された特別なATP駆動ポンプ
- これはP-gp(P-糖タンパク質)として知られるABC輸送体の一種です。
- P-gpは多剤耐性タンパク質として発見され、がん細胞が化学療法薬を排出し耐性を獲得する機構の主役です。
5. P-gpの天然基質だと現在研究者が考えているもの
- これらの多剤排出ABCトランスポーターの天然の基質は、脂質関連分子やステロイド、代謝産物など多様な有機分子であると考えられています。
- つまり、細胞はもともと細胞内の不要物や代謝物、異物を除去するためにこのポンプを利用しており、抗がん剤はこうした輸送系の「誤認識」や「偶然の基質」として排出されると考えられています。
- このポンプの機能ががん細胞の薬剤耐性を生み出す重要因子となっています。
- P-gpは人工薬剤以外に、生体内の脂質やステロール類、脂肪酸誘導体、ペプチドなどの疎水性または両親媒性の自然基質を輸送している可能性が強く示唆されています。
- 例えば、胆汁酸やホスファチジルセリンなど細胞内脂質関連物質の移動に関与し、生理的な膜脂質の恒常性維持に役立っていると考えられています。
まとめ
| 種類 | 輸送対象 | 代表的機能 |
|---|---|---|
| P型ATPアーゼ | 主にイオン | Na⁺/K⁺ポンプ、Ca²⁺ポンプ |
| F型ATP合成酵素 | 主にH⁺イオン(ATP合成) | ATP合成 |
| V型ATPアーゼ | H⁺イオン | リソソームの酸性化 |
| ABC輸送体 | 薬剤や脂質・ペプチドなど多様な小分子 | 多剤排出、脂質輸送 |
- P-gpはABC輸送体で、多剤耐性薬の輸送で発見されたが、天然では主に脂質などの生理的基質を輸送していると考えられている。
このようにATP駆動ポンプは多様な分子輸送と細胞機能維持に寄与しています。
ゲノム解読は続き、ゲノムの塩基配列がすべて明らかになった生物も増えている。こうした情報からマウスあるいはヒトに特定の輸送体またはポンプが何個存在するかを調べるにはどうしたらよいか。塩基配列から同定された輸送体またはポンプの多くは、その天然の基質や生理的役割がわからないという意味で“オーファン”タンパク質である。どうしてそうなるのか。こうした“オーファン”タンパク質の生理的役割を明らかにするにはどうしたらよいのか。
1. 特定の輸送体やポンプの数をゲノム情報から調べる方法
- ゲノムデータベースの利用
マウスやヒトのゲノム配列データは公的データベース(例:NCBI、Ensembl、UCSC Genome Browser)で公開されています。 - バイオインフォマティクス解析
- 既知の輸送体やポンプのタンパク質配列や機能ドメイン情報を使い、「ホモロジー検索(BLASTなど)」で類似遺伝子を検索します。
- 家族・スーパー何の分類が行われているため、例えばABCトランスポーター、チャネル類をそれぞれグループごとに検索・カウント可能。
- タンパク質構造予測やモチーフスキャンも併用し、候補を的確に抽出。
2. “オーファン”タンパク質が多い理由
- ゲノム解読は全ての遺伝子を特定できても、
- どの遺伝子がどんな基質を輸送しているのか(機能)や、
- その細胞内での生理的役割や重要性まではすぐには分かりません。
- また、実験的に研究が進んでいない新規遺伝子や、複雑な調節・相互作用を持つもの、あるいは特定条件下でのみ機能するものも多い。
- これらは「オーファン(孤児)」タンパク質、すなわち「未知の機能を持つタンパク質」と呼ばれます。
3. “オーファン”タンパク質の生理的役割を明らかにする方法
- 発現解析
- どの組織・臓器、発生段階、細胞周期で発現しているかを解析
- RNA-seqやプロテオミクスで発現状況を確認
- 遺伝子改変実験
- ノックアウト(遺伝子破壊)や過剰発現で、細胞または動物レベルで機能や表現型を調べる
- CRISPR-Cas9やRNAi技術が利用される
- 基質特異性解析
- ホモロジー解析から推定される基質との相互作用実験や、輸送活性のin vitro測定
- 蛍光標識基質や放射性物質を用いた輸送実験
- 細胞内局在の解析
- 蛍光染色や免疫染色で、どの細胞内コンパートメントに存在するか調べる
- 相互作用解析
- 他のタンパク質との相互作用ネットワークを明らかにし、機能推定
まとめ
- ゲノムデータとバイオインフォマティクスを駆使し、輸送体・ポンプを網羅的に同定できる。
- しかし、多くはまだ機能不明の「オーファン」
- 発現解析、遺伝子改変、輸送活性測定など多角的アプローチで生理的役割を解明することが重要である。
この研究は新規治療標的や生物学的理解の深化につながります。
特定のチャネルやポンプや輸送体に対する阻害剤や活性化剤は製薬業界が産生する薬のかなり大きな部分を占めている。骨格筋の収縮は細胞質Ca2+濃度上昇によってひき起こされる。筋小胞体(SR)のP型Ca2+ ATPaseを選択的に阻害する薬物は筋収縮にどのような影響を与えると考えられるか。
筋小胞体(SR)P型Ca²⁺ ATPaseの機能
- 筋小胞体(SR)のP型Ca²⁺ ATPase(通称:SERCA)は、筋収縮後に細胞質内のCa²⁺イオンをSR内部に能動的に戻す働きをしています。
- この機構によって、細胞質のCa²⁺濃度が低下し、筋肉が弛緩します。
阻害された場合の筋収縮への影響
- SERCAを選択的に阻害する薬物が作用すると、Ca²⁺がSRに戻されず、筋細胞質内に残り続けるため、
- 細胞質Ca²⁺濃度の高い状態が持続し、筋肉は弛緩しにくく、持続的な収縮状態(収縮亢進)や痙攣(けいれん)が引き起こされる可能性が高くなります。
- 筋肉は“収縮→弛緩→再収縮”というサイクルが必要ですが、弛緩ができなくなり正常な筋運動が阻害される危険があります。
まとめ
筋小胞体のP型Ca²⁺ ATPase(SERCA)を阻害する薬物は、筋弛緩ができなくなり骨格筋の収縮が持続/亢進する(過収縮・痙攣)という作用をもたらします。この仕組みはカルシウムの細胞内濃度制御が筋収縮に直結することを示す典型例です。
植物とは違い、動物細胞の膜電位はおもに静止K⁺チャネルによって生じる。これらのチャネルはどのようにして静止膜電位形成に寄与するのか。なぜこれらのチャネルは開閉調節を受けないとみなされているのか。これらのチャネルはどのようにK⁺とNa⁺を区別しているのか。
1. 静止K⁺チャネルによる静止膜電位形成
- 動物細胞ではK⁺(カリウム)濃度は細胞内が高く、細胞外が低いという特徴があり、静止K⁺チャネル(しずしK⁺チャネル=リークK⁺チャネル)は常に開いていて、K⁺が濃度勾配に従い細胞外へ流れ出します。
- この結果、細胞内に相対的な負電荷が残り、細胞膜の内外に電位差(静止膜電位)が生じます。通常、静止膜電位は-60~-90 mV程度です。
- 静止K⁺チャネルが主に開いていることで、膜のK⁺への透過性が高まり、Na⁺など他のイオンと比べてK⁺が支配的に膜電位を決めています。
2. 開閉調節を受けないとみなされる理由
- 静止K⁺チャネルは一般に「常に開いている」状態で、特別な刺激や膜電位変化による開閉調節機構(ゲート機構)を持たないと考えられています。
- 活動電位を担うNa⁺チャネルや電位依存性K⁺チャネルとは違い、これらは細胞の興奮等による「ゲート調節」をほとんど受けず、安定的に膜電位形成の基盤を供給します。
3. K⁺とNa⁺の選択的透過性
- 静止K⁺チャネルは選択的フィルター構造を有しており、チャネル内部はK⁺イオンのサイズと電荷、脱水和状態などに合わせてK⁺のみ高効率で通過可能な分子構造になっています。
- Na⁺イオンはサイズや水和半径がわずかに異なるため、静止K⁺チャネルの選択的フィルターを通過できないか、効率が極めて低い。このため、K⁺とNa⁺が明確に区別されます。
まとめ
- 静止K⁺チャネルは、K⁺イオンの透過を介して動物細胞の静止膜電位形成に中心的役割を担う。
- これらのチャネルは主に「開いたまま」で、特別な調節を受けない。
- K⁺のみを通す精密な分子選択フィルターにより、Na⁺との識別が実現されている。
パッチクランプ法により個々のイオンチャネルの電気伝導度を測定することができる。K⁺チャネルと思われる遺伝子産物が実際にK⁺あるいは Na⁺チャネルであることを確認する上でパッチクランプ法はどのように使われるかを述べよ。
パッチクランプ法は、細胞膜上の単一または複数のイオンチャネルを通るイオン電流(微小電流)をリアルタイムで測定する強力な手法です。K⁺チャネルとNa⁺チャネルの違いを実際に判別する際には以下の流れで用いられます。
1. イオン選択性の判定
- 遺伝子産物(例:K⁺チャネルcDNA)を発現させた細胞やリポソームにパッチクランプ電極を密着させて単一チャネルまたは全細胞記録を行う(cell-attachedまたはwhole-cellモードなど)。
- 外液や内液のイオン組成(K⁺濃度とNa⁺濃度)を意図的に変化させ、特定イオン(K⁺やNa⁺)の濃度勾配を操作する。
- その状態で、チャネルから観察されるイオン電流の方向や大きさ(コンダクタンス)の変化を測定する。
2. 判定ポイント
- チャネルを通して流れる主なイオンがK⁺であれば、外液・内液のK⁺濃度差によって電流の大きさやNernst電位(平衡電位)が変わります。
- 逆に、Na⁺が主に通るなら、Na⁺濃度勾配に応じて電流が変化し、Na⁺のNernst電位に沿った挙動を示します。
- 実際に外液のK⁺濃度だけを変える実験や、Na⁺のみを高濃度にしてK⁺を極端に少なくするなどしてみることで、どのイオンが優先的にチャネルを通るかを判定できます。
3. 薬理学的検証
- テトロドトキシン(TTX:Na⁺チャネル阻害剤)や四アンモニウム(TEA:K⁺チャネル阻害剤)など、イオンチャネルごとに特異的な阻害剤を加えることで、その影響からチャネルの種類を特定することもできます。
まとめ
パッチクランプ法を用いて、チャネル遺伝子産物の発現細胞で
1.イオン環境を制御しながら電流を測定
2.電流の特性、平衡電位、阻害剤感受性を解析
することで、そのチャネルがK⁺チャネルかNa⁺チャネルかを明確に区別することができます。
これにより遺伝子産物が実際に目的とするイオンチャネルであることを機能的に証明できます。
植物は液胞膜をはさんでのH⁺の電気化学的配を使って液胞に塩や糖をたくわえている。これにより液胞は高張になる。植物細胞はなぜこれによって破裂しないのか。動物細胞膜のN⁺/K⁺ATPase は張条件下での動物細胞の破裂を防いでいる。どのようにしてそれを行っているのか。
植物細胞が高張液胞でも破裂しない理由
- 植物細胞は分厚くて丈夫な「細胞壁」で囲まれているため、
液胞に塩や糖を蓄えて「高張(浸透圧が高い)」状態になっても、細胞内へ水が流入して膨張した際に、細胞壁が物理的に細胞の破裂を防ぎます。 - 液胞に水が入り膨圧(細胞壁を外へ押す力)が高まっても、細胞壁が耐えるため「ちょうどよい硬さ」になり、細胞は破裂せず立体的な形や植物体の硬さ(しおれ防止)も維持できます。
動物細胞膜のNa⁺/K⁺ATPaseによる破裂防止機構
- 動物細胞には細胞壁がないため、細胞膜だけでは膨張に耐えきれず、浸透圧バランスが崩れると破裂(溶血)します。
- Na⁺/K⁺ATPase(ナトリウムポンプ)は、細胞内のNa⁺を積極的に外に出し、K⁺を細胞内に取り込むことで「細胞内の浸透圧を抑え」ています。
- 細胞外Na⁺濃度を高く、細胞内Na⁺濃度を低くして、細胞内への水の流入を防ぎ、浸透圧バランスを保つことで細胞膜の破裂を防いでいるのです。
まとめ
- 植物細胞は頑丈な細胞壁のおかげで、高張な液胞になっても破裂せず、「膨圧」の力で細胞や植物体を支えています。
- 動物細胞はNa⁺/K⁺ATPaseの働きにより、細胞内外のイオンバランスを整え、浸透圧の高まりによる水の流入や破裂を防いでいます。
このため、植物も動物もそれぞれの方法で「高浸透圧ストレス」から細胞を守っているのです。
細菌の Na⁺/ロイシン輸送体の場合、結合イオンをK⁺ではなく Na⁺にしていることの重要な点は何か。小腸上皮細胞においてグルコースを等方輸送するしくみを説明せよ.この輸送に使われるイオンは何で、このイオンが細胞膜を通って流入することがエネルギー的に好都合となる理由を二つあげよ。
1. 細菌Na⁺/ロイシン輸送体においてNa⁺を結合イオンにする重要な理由
- 多くの細菌は細胞外の環境にNa⁺が豊富であり、K⁺より遥かに高濃度にあります。
- 細胞膜はNa⁺の濃度勾配(外>内)が大きいため、Na⁺を使うことで、濃度勾配をエネルギー源としてロイシンを効率よく取り込めるのが重要です。
- K⁺は細胞内外で勾配が乏しく、エネルギー源になりません。よって、Na⁺利用は生理的な環境と輸送効率の両面で合理的です。
2. 小腸上皮細胞でのグルコース等方輸送のしくみ
- 小腸上皮細胞ではNa⁺/グルコース共輸送体(SGLT:Sodium Glucose Linked Transporter)を用いて、グルコースを細胞内に取り込みます。
- 等方輸送=濃度勾配に逆らってグルコースを取り込み可能で、これはNa⁺の濃度勾配に依存した「二次的能動輸送」です。
3. グルコース輸送に使われるイオンとその好都合なエネルギー理由
- この輸送に使われる主なイオンはNa⁺(ナトリウムイオン)です。
- Na⁺が細胞膜を通って流入することがエネルギー的に好都合な理由は:1.Na⁺の濃度勾配(外側が高く、内側が低い)をエネルギー源として利用できること
→ Na⁺/K⁺ATPaseの働きにより、細胞内外でNa⁺勾配が保たれているので、Na⁺が自発的に細胞内に流れ込める。このポテンシャル差を利用して、グルコースも一緒に取り込むことができる。2.膜電位差(細胞内外の電気的ポテンシャル差)が加算的なエネルギー源になること
→ 細胞外は陽性、細胞内は陰性なので、Na⁺は電気的にも流入しやすく、グルコースの輸送に更なるエネルギーが供給される。
まとめ
- 細菌ロイシン輸送体は外部Na⁺利用で濃度勾配を最大限活用できる。
- 小腸SGLTはNa⁺とグルコースの共輸送を使い、Na⁺勾配と膜電位差の2つのエネルギー源でグルコースを積極的に吸収できる。
このしくみが、効率の良い養分取り込みと生命活動の基盤となっています。
ゼブラフィッシュなどいくつかの生物のゲノム塩基配列が解明され、多数の受容体や輸送体の存在が明らかになった。ゼブラフィッシュのgolden 遺伝子をポジショナルクローニングによって調べたところ、陽イオン交換タンパク質と思われる SLC24A5であることがわかった。このgoldlen遺伝子に突然変異が起こると、メラニンという黒色色素の量が大きく減るので、通常水平方向に走っている黒い縞が薄くなり、全体が金色に見えるようになる、魚類やヒトにあるメラニンを含む膜小胞は何とよばれるか。このgolden 遺伝子からつくられるタンパク質がゼブラフィッシュのどこで発現されどこに局在するのかを明らかにする実験計画を立案せよ。このゼブラフィッシュのgolden 遺伝子に突然変異が起こったときに、他の動物のオルソロガス遺伝子を発現させて、その表現型をもとに戻すことを何というか。
1. メラニンを含む膜小胞の名称
- メラニン色素を含む膜小胞はメラノソームと呼ばれています。
- メラノソームは色素細胞(メラノサイト)内でメラニンを合成・蓄積・輸送するオルガネラです。
2. golden遺伝子から作られるタンパク質の発現・局在を明らかにする実験計画
実験目的
ゼブラフィッシュのgolden遺伝子(SLC24A5)がどの組織で発現し、細胞内のどのコンパートメントに局在するかを明らかにする。
実験方法
- mRNA発現解析
- in situハイブリダイゼーションによりゼブラフィッシュ胚や組織切片中でgolden遺伝子のmRNAを検出。発現細胞や組織を特定。
- qRT-PCRにて組織別発現量を定量。 - タンパク質局在の解析
- golden遺伝子産物に特異的な抗体を用いた免疫染色・免疫蛍光染色を実施。
- メラノソームや細胞内小胞マーカー(例えばLAMP1など)との共染色により、タンパク質の細胞内局在を判定。
- GFPタグを融合したgoldenタンパク質の遺伝子を用いて生きた細胞や胚でライブイメージングも可能。 - 遺伝子編集や変異体解析
- golden突然変異体と正常体で発現・局在の違いを比較。
3. 他動物のオルソログ遺伝子で表現型を回復させることの名称
- 目的の遺伝子が機能しない突然変異体に、同じ機能を持つ他動物由来のオルソログ遺伝子を導入し、表現型を回復させることを「遺伝子補完(Complementation)」と言います。
まとめ
| 項目 | 回答 |
|---|---|
| メラニン含有膜小胞の名称 | メラノソーム |
| golden遺伝子発現・局在解析法 | in situハイブリダイゼーション、免疫染色+共染色、GFPタグによるライブイメージング |
| 他生物オルソログ遺伝子導入による表現型回復 | 遺伝子補完(Complementation) |
このようにして、Golden遺伝子の機能解析とメラノソームにおける役割を詳しく調べることができます。
小腸上皮細胞での内腔側から血液側へのグルコース輸送は経細胞輸送の重要な例である。Na⁺/K⁺ATPase はどのようにこの過程の原動力となっているのか。この過程に密着結合が重要なのはなぜか。スポーツドリンクのような水分補給のための飲料は糖と塩を含んでいる。なぜその二つが水分補給に重要なのか。
1. Na⁺/K⁺ATPaseがグルコース輸送の原動力となる仕組み
- 小腸上皮細胞の基底側(血液側)膜にはNa⁺/K⁺ATPaseがあり、これが細胞内のNa⁺濃度を低く維持しています。
- その結果、細胞の内腔側(小腸内)との間には大きなNa⁺濃度勾配が形成され、Na⁺は内腔側から細胞内へ流れ込みやすくなっています。
- Na⁺/グルコース共輸送体(SGLT)が内腔側膜にあり、このNa⁺濃度勾配を利用して、Na⁺とともにグルコースを能動的に細胞内に取り込みます。
- つまり、Na⁺/K⁺ATPaseが作るNa⁺濃度差が、グルコース輸送のエネルギー源となっています。
2. なぜこの過程に密着結合が重要か?
- 小腸上皮細胞は互いに密着結合(tight junction)で結ばれており、細胞間隙を通っての物質の漏れを防ぎます。
- このため物質は細胞を「経由」して移動(経細胞輸送)することになり、輸送機構の制御が明確になります。
- もし密着結合がなければ、物質は経細胞間隙(細胞間輸送)を通ってしまい、腸管での選択的な吸収ができません。
3. なぜスポーツドリンクに糖と塩を含めると水分補給に効果的か?
効率的な水分補給のカギは、小腸での「ナトリウムとグルコースの共輸送(SGLT)」にあります。
単なる水を飲むよりも、糖(グルコース)と塩分(ナトリウム)を同時に摂取する方が、以下のメカニズムによって格段に吸収スピードが上がります。
- 糖と塩の相乗効果: 小腸の細胞にある輸送体(SGLT1)は、ナトリウムとグルコースがセットになることで初めて働きます。糖を一緒に摂ることで、ナトリウムの吸収が劇的に加速します。
- 水を引き込む「浸透圧」の力: ナトリウムとグルコースが細胞内に吸収されると、細胞内や血液側の溶質濃度(浸透圧)が高まります。すると、体は濃度を一定に保とうとして、腸管内の水を血液側へと強力に引き込みます。
つまり、糖と塩は単なる栄養補給ではなく、水を体内に流し込むための「呼び水」としての役割を果たしているのです。これにより、脱水状態からの迅速な回復が可能になります。
まとめ
| 項目 | 内容 |
|---|---|
| Na⁺/K⁺ATPaseの役割 | 基底膜でNa⁺を排出し、細胞内のNa⁺濃度を低く保つことで、グルコース輸送のエネルギー源(濃度勾配)を作る。 |
| 密着結合の役割 | 細胞間隙を密閉して物質の漏れを防ぎ、必要な物質を細胞経由で確実に運ぶ(経細胞輸送)ためのバリアとなる。 |
| 糖と塩の重要性 | 糖がNa⁺の効率的な吸収を促し、それによって生じる浸透圧の変化が水を強力に引き込む。この相乗効果で水分補給の効率が最大化される。 |
この仕組みを理解すると、水分補給や栄養吸収の仕組みがよくわかります。
問題文引用元:東京化学同人 分子細胞生物学 第6版
コメント