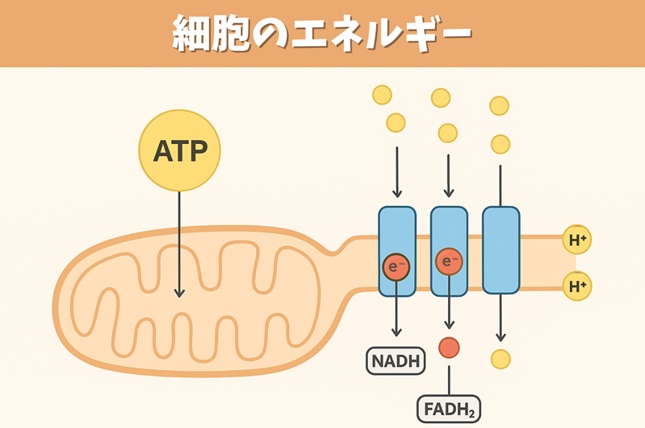
- プロトン駆動力(pmf)はミトコンドリアと葉緑体の機能に必須なものである。何がpmfをつくり出し,そしてpmfはATPとどう関係するのか。
- ミトコンドリア内膜は典型的細胞膜の性質をすべてもっているが、そのほかに酸化的リン酸化を行うための特別な性質をもっている。その特別な性質とは何か。それらはどのように内膜の機能とかかわっているのか。
- グルコースから最大量のATP をつくるためには解糖系、クエン酸回路,電子伝達系での反応が必要である。これらのうちでO2を必要とするものはどれか。それはなぜか。これらのうちのどれが、ある種の生物の中あるいはある種の生理的条件下で、O2なしで進行できるか。
- 解糖系で生じた電子がどのように電子伝達鎖に運ばれるかを説明せよ。この輸送系が働かなくなるような突然変異が起こったとき、グルコース 1分子からつくられる ATP量はどうなるか。この状態が長引くと解糖系にはどのような影響がでるか。
- ミトコンドリアでの脂肪酸の酸化は主要な ATPの供給源であるが、脂肪酸の酸化は他の場所でも起こっている。ミトコンドリア以外で脂肪酸を酸化できる細胞小器官は何か。その細胞小器官での酸化とミトコンドリアでの酸化で大きく異なる点は何か。
- ミトコンドリアのシトクロムはそれぞれ補欠分子族をもっている。補欠分子族とは何か。シトクロムに結合しているものはどのようなグループに属するものか。電子伝達鎖中を電子が一方向に伝達されていくのは種々のシトクロムのどのような性質によるのか
- NADHから1対の電子が伝達されると約3分子のATPが合成されるが、FADHからだと約2分子しか合成されない。この電子対当たりの ATP合成量の違いは何によるのか
- ATP 合成酵素に関する知識の多くは好気性細菌の研究によって得られた。この生物のどのような点が研究に有利なのか。この生物の解糖系,クエン酸回路、電子伝達鎖は細胞のどこで行われているのか。好気性細菌のpmfはどこに形成されるのか。これらの生物において、pmfは他のどのような細胞活動に使われるのか
- ミトコンドリア内膜の重要な機能の一つは選択的透過性によって水溶性分子の透過を制限し、膜の内外で異なった化学環境をつくることである。しかし、酸化的リン酸化の基質や生成物の多くは水溶性であるのに内膜を通り抜けねばならない。この輸送はどのように行われているのか
- ミトコンドリアと葉緑体と細菌の電子伝達鎖においてQ回路は重要な役割を果たしている。Q回路の機能は何か。それはどのように行われているのか。ミトコンドリアや紅色細菌や葉緑体のQ回路において電子伝達を行っている構成成分は何か
- 光合成における酸素発生の反応式を書け。“光合成における酸素の発生は炭水化物やATP をつくる反応の副産物にすぎない”という文の意味を説明せよ
- 光合成は複数の過程に分けることができる。光合成にはどのような過程があり、それらは葉緑体のどこで行われるのか。光合成においてスクロースはどこでつくられるのか
- 光のエネルギーを吸収する光化学系は反応中心とアンテナ複合体という二つの密着した成分からなる。それぞれの色素成分と、それらが光吸収過程で果たす役割は何か。それぞれに含まれている色素が光合成に関与していることを示す証拠にはどのようなものがあるか。
- 緑色および紅色細菌の光合成ではO2は発生しない。なぜか。これらの生物が光合成を使ってATPを合成できるのはなぜか.これらの生物では何が電子供与体となっているのか
- 葉緑体には二つの光化学系が存在する。それぞれの機能は何か。線形電子伝達で、光吸収から NADPH 生成までの電子の流れを図で示せ。NADPHにたくわえられたエネルギーから何がつくられるのか
- 暗所ではCO2を固定するカルビン回路の反応は行われない。その理由として何が考えられるか。これらの反応はどのようにして光によって調節されるのか
- 地球上に最も大量に存在するタンパク質であるrubiscoは光合成生物における炭水化物合成に重要な役割を果たす。rubiscoとは何で、細胞内のどこに存在し、どのような機能を果たしているのか
プロトン駆動力(pmf)はミトコンドリアと葉緑体の機能に必須なものである。何がpmfをつくり出し,そしてpmfはATPとどう関係するのか。
pmfをつくり出すもの
- ミトコンドリアでは、内膜の電子伝達系(複合体I、III、IV)が基質の酸化で得た自由エネルギーを用いてH⁺をマトリックスから膜間腔へ汲み出し、膜を挟む電気化学的なH⁺勾配を形成する。ここでの勾配は膜電位成分(Δψ)とpH勾配成分(ΔpH)の組み合わせで定義される。
- 葉緑体では、チラコイド膜の光化学系(光合成電子伝達)が水の酸化から得たエネルギーでストロマからルーメンへH⁺を蓄え、主にΔpHに富むプロトン駆動力を形成する(Δψはイオン移動で部分的に緩和されやすい)。
pmfとATPの関係
- pmfはF型ATP合成酵素(F₀F₁-ATP synthase)の「回転モーター」を駆動する原動力である。H⁺が濃度・電位勾配に従ってF₀部分を通過すると回転トルクが生じ、その機械的エネルギーがF₁部分の化学反応中心に伝達され、ADPと無機リン酸(Pi)からATPが合成される。
- したがって、電子伝達系(呼吸鎖あるいは光リン酸化)が「pmfをつくる工程」、ATP合成酵素が「pmfをATPに変換する工程」を担い、化学浸透説に基づくエネルギー変換が完結する。
ミトコンドリア内膜は典型的細胞膜の性質をすべてもっているが、そのほかに酸化的リン酸化を行うための特別な性質をもっている。その特別な性質とは何か。それらはどのように内膜の機能とかかわっているのか。
特別な性質の要点
- 非常に低い透過性と選択的トランスポーターの豊富さ:内膜は水やイオンに対してほぼ不透過で、特定基質用のキャリア(ADP/ATPトランスロカーゼ、リン酸キャリア、ピルビン酸キャリアなど)によってのみ交換・輸送が起こる。
- クリステ構造による巨大な表面積と区画化:内膜はクリステ(ひだ)により表面積を大幅に増大し、呼吸鎖複合体とATP合成酵素を高密度に配置できるようにしている。
- カルジオリピンの高含量:内膜に特有のリン脂質カルジオリピンが多く、呼吸鎖超複合体の安定化やプロトン保持に寄与する。
- 呼吸鎖複合体とATP合成酵素の組織化:複合体I–IVとATPシンターゼがクリステに沿って配置・会合し、効率的な電子移動とプロトン勾配の形成・利用を可能にする。
- クリステネック(クリスタジャンクション)とMICOSによるドメイン分離:内膜は内側境界膜とクリステ膜に機能分化し、狭いネック構造と内膜タンパク質複合体(MICOS)で組織化される。
機能との関わり
- プロトン駆動力(pmf)の生成と保持:低透過性とカルジオリピン、クリステ形態により、呼吸鎖が汲み出したH⁺を膜間腔側に効率よく蓄積し、ΔψとΔpHからなるpmfを強固に維持する。
- ATP合成の高効率化:クリステで表面積・曲率を最適化し、呼吸鎖とATPシンターゼの距離を近接・配列化することで、H⁺のリークを最小化しつつF0F1-ATP合成酵素の回転触媒を最大効率で駆動する。
- 代謝物流の選択的制御:豊富な専用キャリアにより、ADP/ATP、Pi、ピルビン酸、アスパラギン酸/グルタミン酸などの交換を精密制御し、酸化的リン酸化と基質供給を緊密に連結する。
- 構造—機能相関の動的調節:MICOSやATPシンターゼのダイマー化がクリステ形態を規定し、細胞のエネルギー需要に応じて内膜の構築を可塑的に変化させ、ATP生産能力を最適化する。
ひと言で
ミトコンドリア内膜は「漏れない・選ぶ・畳む・組む」という特別な性質(超低透過性、選択的キャリア、クリステ拡大、呼吸鎖とATPシンターゼの組織化)によって、プロトン駆動力を高効率で生み・保持し・ATPへ変換するための専用プラットフォームとして機能している。
グルコースから最大量のATP をつくるためには解糖系、クエン酸回路,電子伝達系での反応が必要である。これらのうちでO2を必要とするものはどれか。それはなぜか。これらのうちのどれが、ある種の生物の中あるいはある種の生理的条件下で、O2なしで進行できるか。
O2を必要とする経路はどれか
- 電子伝達系(呼吸鎖)はO2を必要とする。複合体IVで最終電子受容体としてO2が還元されてH2Oが生成し、この反応が電子の流れとプロトン汲み出しを駆動してプロトン駆動力を形成するため、O2が欠けると電子伝達が停止する。
- クエン酸回路と解糖系そのものは反応式にO2を直接使わないが、クエン酸回路で生じるNADHやFADH2の再酸化は通常電子伝達系に依存し、その電子伝達系がO2を要するため、好気条件下での連続運転にはO2が実質的に不可欠となる。
O2なしで進行できるものは何か
- 解糖系はO2なしでも進行できる。嫌気条件では、ピルビン酸を乳酸(乳酸発酵)やエタノールとCO2(アルコール発酵)に還元してNAD+を再生し、解糖を継続できる。
- 一部の微生物や特殊条件では、クエン酸回路の全体ではなく部分経路が回り、還元力や前駆体供給のために用いられることがある。だが多くの動物細胞では、O2がないとNADH再酸化が滞り、クエン酸回路は実質的に停止する。
- 電子伝達系は好気呼吸ではO2が必須だが、嫌気呼吸を行う微生物は硝酸イオンなど別の最終電子受容体を用いてO2なしで電子伝達を回すことがある(動物細胞には基本的に備わっていない)。
まとめの対応関係
- O2が必要:電子伝達系(好気呼吸の最終電子受容体がO2であるため)
- O2がなくても可能:解糖系(発酵でNAD+再生を伴えば継続可能)
- 条件次第で一部継続:クエン酸回路(動物細胞では実質停止、微生物では代替受容体や部分運転の例あり)。
解糖系で生じた電子がどのように電子伝達鎖に運ばれるかを説明せよ。この輸送系が働かなくなるような突然変異が起こったとき、グルコース 1分子からつくられる ATP量はどうなるか。この状態が長引くと解糖系にはどのような影響がでるか。
電子はどう運ばれるか
輸送系が働かない突然変異でのATP収量
- シャトルが機能しないと、細胞質のNADHは電子伝達系に供給できず、酸化的リン酸化によるATP産生が低下する。
- 解糖自体の基質レベルリン酸化は残るため、グルコース1分子あたりのATPは「解糖の純利得」中心にほぼ限定され、好気条件の最大収量より大幅に減少する(理論的には2 ATP程度に近づく)。
- グリセロールリン酸やマレート–アスパラギン酸の経路が完全に途絶すると、解糖由来NADHは呼吸鎖に入れず、ミトコンドリアでのATP生成分が失われる。
長引くと解糖系に出る影響
- 細胞質NADHが再酸化されずNAD+が枯渇し、グリセルアルデヒド-3-リン酸脱水素酵素反応が停滞して解糖フラックスが落ちる。
- 代償的に乳酸発酵などでNAD+再生が亢進し、ピルビン酸→乳酸が増えて細胞外酸性化や乳酸蓄積が起こる(好気的条件でも擬似的に発酵依存に傾く)。
- その結果、ATP供給は主に基質レベルリン酸化に依存し、全体のエネルギー産生能力が低下、長期的には代謝リモデリングや機能不全につながる。
ミトコンドリアでの脂肪酸の酸化は主要な ATPの供給源であるが、脂肪酸の酸化は他の場所でも起こっている。ミトコンドリア以外で脂肪酸を酸化できる細胞小器官は何か。その細胞小器官での酸化とミトコンドリアでの酸化で大きく異なる点は何か。
どの細胞小器官か
- ミトコンドリア以外で脂肪酸の酸化を行う細胞小器官はペルオキシソームである。
大きく異なる点
- 酵素系と目的の違い
- ペルオキシソームのβ酸化は、極長鎖脂肪酸(C20以上)や分枝鎖・非常に長い脂肪酸の鎖短縮に特化し、主に「前処理(短鎖化)」を担う。できた中鎖〜短鎖のアシルCoAやアセチルCoAはしばしばミトコンドリアへ送られ、そこで完全酸化される。
- 初段の電子受容体の違い
- ミトコンドリアのβ酸化は脱水素で生じたFADH2やNADHを呼吸鎖に渡してATP合成に結びつける。
- ペルオキシソームの初段酸化はフラビン酸化酵素から電子が直接O2へ渡され、過酸化水素(H2O2)が生じる。H2O2はカタラーゼで分解され、ここでは電子伝達鎖を介したATP産生に直結しない。
- エネルギー収支
- ミトコンドリアのβ酸化は生じた還元当量を電子伝達系に供給して酸化的リン酸化により多量のATPを得る。
- ペルオキシソームのβ酸化はATP産生にほとんど直接寄与せず、鎖長短縮と解毒(H2O2分解)が中心で、エネルギー効率は低い。
- 基質選好性と輸送
- ミトコンドリアは主に長鎖脂肪酸(C16–C18)をカルニチンシャトルで取り込み、反復β酸化で完全酸化する。
- ペルオキシソームは極長鎖脂肪酸や特殊脂肪酸を取り込み、部分酸化してから産物を他オルガネラ(多くはミトコンドリア)へ受け渡す。
- 連携
- ペルオキシソームでの鎖短縮後、ミトコンドリアでの完全酸化という分業で、難処理の脂肪酸も最終的にエネルギー化できる。
要するに、ペルオキシソームは「特殊・極長鎖脂肪酸の前処理とH2O2処理」を担い、ミトコンドリアは「還元当量をATPに変換しつつ完全酸化」を担うという役割分担が、脂肪酸酸化の大きな違いである。
ミトコンドリアのシトクロムはそれぞれ補欠分子族をもっている。補欠分子族とは何か。シトクロムに結合しているものはどのようなグループに属するものか。電子伝達鎖中を電子が一方向に伝達されていくのは種々のシトクロムのどのような性質によるのか
補欠分子族とは
補欠分子族は、タンパク質本体(ポリペプチド鎖)に強く結合して機能発現に必須の役割を果たす非タンパク質性の成分で、金属錯体や有機補酵素などが該当する。酵素や電子伝達タンパク質では、この補欠分子族が酸化還元や基質結合の実体として機能し、タンパク質はその化学中心を適切な環境に配置・制御する役目を担う。
シトクロムに結合している補欠分子族のグループ
シトクロムはヘム(鉄ポルフィリン錯体)を補欠分子族として持つヘムタンパク質であり、ヘムの種類に応じてシトクロムa、b、cなどに分類される。いずれもヘム鉄の原子価(Fe2+/Fe3+)の可逆変化によって電子の受け渡しを行うのが本質で、ヘモグロビンのようにO2結合を主機能とするヘムタンパク質とは機能の中心が異なる。ミトコンドリア電子伝達鎖では、複合体III(シトクロムbc1複合体)にb型・c1型のヘム、可溶性のシトクロムcにc型ヘム、複合体IV(シトクロムcオキシダーゼ)にa型ヘム(ヘムa、ヘムa3)と銅中心(CuA、CuB)が配置され、段階的な電子移動を担う。
電子が一方向に伝達される理由(性質)
電子が一方向へと流れるのは、電子伝達体間の酸化還元電位(還元電位、E°’)の段階的勾配が設計されているためである。一般に、電子はより低い酸化還元電位のキャリアから、より高い酸化還元電位のキャリアへ自発的に移動する。ミトコンドリア内膜に並ぶ各シトクロムのヘム環境(リガンド、周囲のアミノ酸場、疎水性環境、ヘムの種類)によって鉄中心の酸化還元電位が微調整され、複合体IIIのヘム群→可溶性シトクロムc→複合体IV(ヘムa→ヘムa3・CuB中心)→O2という順に、熱力学的に有利な電子移動が起こるように段差が設定されている。さらに、タンパク質間の立体配置・結合部位の特異性と近接距離(電子トンネルの許容距離)、反応速度論的整合(前後段の会合・解離速度、内部再配向)も組み合わさり、逆向きの電子戻りが抑えられて一方向性が実現される。
要点の整理
- 補欠分子族:タンパク質に結合して機能の実体を担う非タンパク質性成分(電子伝達では酸化還元中心)。
- シトクロムの補欠分子族:ヘム(鉄ポルフィリン)。ヘムの種類とタンパク質環境によりシトクロムa/b/cなどに分類。
- 一方向性の根拠:シトクロム間の酸化還元電位が段階的に高くなるよう調整され、立体配置・距離・会合解離の速度論的要因と相まって、電子は熱力学的に有利な方向へ連続的に流れる。
NADHから1対の電子が伝達されると約3分子のATPが合成されるが、FADHからだと約2分子しか合成されない。この電子対当たりの ATP合成量の違いは何によるのか
NADHは複合体Iから電子を供与して複合体I・III・IVの3カ所でプロトンポンプを駆動するが、FADH2は複合体IIから入り複合体Iをバイパスするため汲み出されるプロトン数が少なく、結果としてATP合成量が小さくなる。
仕組み
- NADHの2電子(1対)あたりに膜間腔へ汲み出されるプロトン数は合計10個(複合体Iで4個、複合体IIIで4個、複合体IVで2個)で、これが約3 ATPに相当する。
- FADH2の2電子(1対)あたりのプロトン数は合計6個(複合体IIIで4個、複合体IVで2個。複合体IIは0個)で、これが約2 ATPに相当する。
根拠と内訳:
- NADHは複合体I→III→IVを経由し、それぞれのプロトンポンプ活性により4+4+2=10 H+が汲み出される。
- FADH2は複合体IIから入り、IIはポンプ活性がないためIIIとIVのみで4+2=6 H+が汲み出される。
- 一般的換算では、約4 H+の逆流でATP合成1分子(F0F1の回転とリン酸輸送等を含む実効値)とされ、10 H+≈約3 ATP、6 H+≈約2 ATPとなる。
要点は「エントリーポイントの違い」と「それに伴うプロトン汲み出し数の差」。複合体Iを経由するNADHはI・III・IVの3段でH⁺ポンピングに寄与するのに対し、複合体IIから入るFADH2はIII・IVの2段のみで寄与するため、ATP収量が少なくなる。
ATP 合成酵素に関する知識の多くは好気性細菌の研究によって得られた。この生物のどのような点が研究に有利なのか。この生物の解糖系,クエン酸回路、電子伝達鎖は細胞のどこで行われているのか。好気性細菌のpmfはどこに形成されるのか。これらの生物において、pmfは他のどのような細胞活動に使われるのか
結論(短答): 好気性細菌は小型・増殖が速く、遺伝学と生化学操作が容易で膜系が単純なためATP合成酵素研究に最適である。 解糖系は細胞質、クエン酸回路は細胞質(原核ではマトリックスに相当する区画なし)だが酵素は細胞質側、電子伝達鎖とATP合成酵素は原形質膜上で機能する。 細菌のpmfは原形質膜(細胞膜)を挟んで形成され、同じ膜上のF0F1-ATP合成酵素でATP合成に使われるほか、べん毛モーター回転、膜輸送(取り込み・排出)、能動輸送体の駆動などにも利用される。
研究に有利な点
代謝の局在(好気性細菌)
pmfの形成場所と用途
要するに、好気性細菌では原形質膜が「ミトコンドリア内膜の機能」を一手に担い、そこで生じるpmfがATP合成のみならず運動や膜輸送のエネルギー通貨として広く使われている。
ミトコンドリア内膜の重要な機能の一つは選択的透過性によって水溶性分子の透過を制限し、膜の内外で異なった化学環境をつくることである。しかし、酸化的リン酸化の基質や生成物の多くは水溶性であるのに内膜を通り抜けねばならない。この輸送はどのように行われているのか
結論(短答): ミトコンドリア内膜は水溶性分子に対して低透過性だが、専用の膜内輸送タンパク質(キャリア/トランスロカーゼ)が基質特異的に交換・共輸送を行うことで、酸化的リン酸化に必要な水溶性基質・生成物を選択的に通過させている。
何が通され、どう運ばれるか
- ADPとATPの交換: ADP/ATPトランスロカーゼ(ANT)が内膜を介して1対1の逆向き交換を行い、マトリックスへADPを取り込みつつATPを細胞質側へ放出する(電位依存の反ポート)。
- 無機リン酸(Pi)取り込み: Pi/H+シンポーター(リン酸キャリア)がプロトン勾配(pmf)のΔpHを利用してPiをマトリックスへ導入し、ATP合成に供給する。
- ピルビン酸取り込み: ピルビン酸/H+シンポーターが解糖産物ピルビン酸を内膜越しに取り込み、ピルビン酸デヒドロゲナーゼ系へ渡す。
- 代謝中間体の交換: リンゴ酸、アスパラギン酸、クエン酸、アデニンヌクレオチド以外の有機酸などは、専用の二塩基性酸キャリア/アスパラギン酸-グルタミン酸キャリア等で電荷・化学量論に応じた交換輸送(アンチポート)でやり取りされ、NADH等価の移送(マレート-アスパラギン酸シャトル)や前駆体供給に寄与する。
- 無機イオン: Pi以外にもCa2+はミトコンドリアCa2+ユニポーター(MCU)複合体で取り込まれ、Na+/Ca2+交換体などで排出される(酸化的リン酸化の調節に関与)。
なぜ選択的透過性が保てるか
- 内膜は極めて低い非特異的透過性(カルジオリピンに富む)を持ち、拡散漏れを抑制するため、基質移動は原則として専用キャリア経由に限定される。これにより膜電位(Δψ)とpH差(ΔpH)から成るプロトン駆動力(pmf)を保持したまま、必要分子だけを能率的にやり取りできる。
- 多くのキャリアは電荷選択性や化学量論(1:1交換、H+共輸送)を備え、輸送そのものがpmfと熱力学的に整合するよう設計されている(例:ANTは陰性のマトリックス側へ正味電荷が大きいATPの移動を避け、ADP/ATPの電位差を利用するなど)。
全体像(酸化的リン酸化との結びつき)
- 電子伝達系が作るpmfはATP合成酵素によりATPへ変換されるが、合成のためのADP・Pi供給と、生成したATPの外への搬出は、内膜キャリア群の協調に依存する。
- 同時に、解糖とTCA・電子伝達をつなぐ炭素・還元当量のやり取り(ピルビン酸、マレート/アスパラギン酸など)も専用キャリアで実現し、内外の化学環境差を維持したまま代謝フラックスを円滑化している。
要点: 低リークな内膜+基質特異的キャリア(交換・共輸送)により、pmfを保ちつつADP/ATP・Pi・ピルビン酸・有機酸などの水溶性分子を選択的に通過させ、酸化的リン酸化を成立させている。
ミトコンドリアと葉緑体と細菌の電子伝達鎖においてQ回路は重要な役割を果たしている。Q回路の機能は何か。それはどのように行われているのか。ミトコンドリアや紅色細菌や葉緑体のQ回路において電子伝達を行っている構成成分は何か
結論(短答): Q回路の機能は、キノール(QH2)の2電子を「1電子×2本の経路」に分配しつつ、膜の向こう側へプロトンを放出してプロトン駆動力(pmf)を強化することにある。これはキノン結合部位(外側Qoと内側Qi)をもつチトクロムbc1(植物ではb6f)複合体で、bヘム鎖とc系電子担体に電子を振り分けることで達成される。ミトコンドリアではユビキノン/ユビキノールとチトクロムb・c1・可溶性シトクロムc、葉緑体ではプラストキノン/プラストキノールとチトクロムb6f・プラストシアニン、紅色細菌ではユビキノン/ユビキノールとbc1・c型シトクロム(あるいはHiPIP)が主要構成要素で電子伝達を担う。
Q回路の機能
- 2電子担体のキノール(QH2)から取り出した2電子を、1本は高電位経路(c系担体)へ、もう1本は低電位経路(bヘム連鎖→Qi部位へ)へ振り分ける「電子分割」により、同時に2個のプロトンを膜外側(ミトコンドリアでは膜間腔、葉緑体ではルーメン)へ放出することでΔpHとΔψを増大させ、pmfを強力に生み出す(プロトンポンプ等価の作用)[概念]。
- その結果、同一のQH2酸化イベントで、電子移動とプロトン放出が結びつき、ATP合成に有利なプロトン勾配が効率的に形成される[概念]。
どのように行われるか(典型的機構)
- 複合体の外側Qo部位でQH2が酸化され、2電子のうち1電子はリースゲ鉄硫黄(Rieske Fe-S)→シトクロムc1→可溶性c型シトクロム(またはプラストシアニン)へ渡る(高電位経路)[概念]。
- 残る1電子はシトクロムbのヘムbL→ヘムbHを通って内側Qi部位のキノンに渡され、Qi部位のQが半還元体セミキノン(- Q−)を形成する(低電位経路)[概念]。
- 次のQH2酸化サイクルで再び同様の分割が起こり、Qi部位のセミキノンは2回目の電子とマトリックス/ストロマ側から取り込む2H+でQH2に還元される。一方、QoでのQH2酸化ごとに2H+が膜外側へ放出されるため、総じて「電子2個の通過で外側へ2H+放出+内側から2H+取り込み」というプロトン循環が成立し、pmfが強化される[概念]。
系ごとの主要構成成分
- ミトコンドリア(呼吸鎖のQ回路)
- キノン系:ユビキノン(Q)/ユビキノール(QH2)
- 複合体:シトクロムbc1(複合体III;Qo/Qi部位、Rieske Fe-S、シトクロムbのヘムbL・bH、シトクロムc1)
- 可溶性キャリア:シトクロムc(インターメンブレンスペース側)
- 葉緑体(チラコイドのQ回路)
- キノン系:プラストキノン(PQ)/プラストキノール(PQH2)
- 複合体:シトクロムb6f(Qo/Qiに相当、Rieske Fe-S、ヘムbL・bH、ヘムf、補助補欠分子であるヘムciなど)
- 可溶性キャリア:プラストシアニン(銅タンパク;ルーメン側)
- 紅色細菌(循環型光合成のQ回路)
- キノン系:ユビキノン/ユビキノール(種によりメナキノンなどの場合も)
- 複合体:チトクロムbc1(Qo/Qi、Rieske Fe-S、ヘムbL・bH、シトクロムc1)
- 可溶性キャリア:c型シトクロム(シトクロムc2)やHiPIP(高電位鉄硫黄タンパク)
光合成における酸素発生の反応式を書け。“光合成における酸素の発生は炭水化物やATP をつくる反応の副産物にすぎない”という文の意味を説明せよ
ポイントの整理
- 目的:2電子担体QH2のエネルギーを最大活用し、1電子キャリア列へ安全に分配しつつ、余剰自由エネルギーをプロトン移動に結び付けてpmfを増強する。
- 手段:Qo/Qiの二座での「分岐酸化」と「セミキノン循環」により、bヘム鎖とc系担体に電子を配分。
- 結果:電子の通過に伴う外側へのH+放出と内側からのH+取り込みが起き、膜を挟んだプロトン勾配(ΔpH、Δψ)が形成・強化される。
このQ回路により、呼吸鎖・光合成鎖の中核でプロトン駆動力が効率良く生成され、下流のATP合成酵素が高効率に駆動される。
光合成における酸素発生の反応式
\[
2\mathrm{H}_2\mathrm{O} \rightarrow \mathrm{O}_2 + 4\mathrm{H}^+ + 4e^-
\]
この反応は光化学系IIで行われ、水分子が光エネルギーにより分解されて酸素分子、プロトン、水素イオン、電子に変換される。
「酸素の発生は副産物にすぎない」という文の意味
- 水の分解は光合成の電子供給源として必要な工程である。水から電子が取り出されることで、電子伝達系を駆動し、ATPやNADPHの合成に使われる電子やプロトンを供給している。
- これらのATPやNADPHはカルビン回路でCO2を還元し、炭水化物を合成するためのエネルギー源および還元力である。
- つまり、光合成の本来の目的は炭水化物やATPの生成であり、酸素の発生は水分解に伴う「不要な産物」あるいは「副産物」にすぎないということを意味する。
- 酸素自体は光合成における主目的ではなく、水の酸化に伴い発生するものとして副次的に生じているのだ。
この理解によって、酸素発生は生命にとって重要な現象でありつつも、光合成のエネルギー変換系の副産物的機能という位置づけになります。
光合成は複数の過程に分けることができる。光合成にはどのような過程があり、それらは葉緑体のどこで行われるのか。光合成においてスクロースはどこでつくられるのか
光合成の主な過程とその場所
- 光化学反応(明反応)
- 葉緑体のチラコイド膜上で行われる。
- 光エネルギーを利用し、水を分解して電子・プロトンを生じさせ、電子伝達系を通じてATPとNADPHを生成する。
- 光化学系II(PSII)と光化学系I(PSI)が連携して電子伝達を行う。
- カルビン・ベンソン回路(暗反応)
- 葉緑体の**ストロマ(液胞様の基質)**内で行われる。
- ATPとNADPHのエネルギーを使い、CO2を固定して3-ホスホグリセリン酸(3-PGA)を経てグルコースなどの炭水化物を合成する。
スクロースの合成場所
- 光合成産物であるグルコースや他の炭水化物は、まずストロマで合成・変換される。
- 合成された糖は細胞の他部位(例:細胞質)の代謝を経て、スクロースは主に葉緑体外の細胞質で合成される。
- スクロースは植物体内輸送の主要な形態で、葉から根などへ運ばれる。
まとめると、光合成は
- チラコイド膜での光化学反応(ATPとNADPH合成)
- ストロマでのカルビン回路(炭水化物合成)
に大別され、
スクロースは葉緑体外の細胞質で作られ、植物体内でエネルギーや炭素の主要輸送体として機能します。
光のエネルギーを吸収する光化学系は反応中心とアンテナ複合体という二つの密着した成分からなる。それぞれの色素成分と、それらが光吸収過程で果たす役割は何か。それぞれに含まれている色素が光合成に関与していることを示す証拠にはどのようなものがあるか。
1. 反応中心の色素成分と役割
- 反応中心は主にクロロフィルaを含む。
- ここに集められた光エネルギーはクロロフィルaの電子を励起し、電子伝達鎖へ電子を渡す反応の起点となる。
- 反応中心クロロフィルaは、電子の放出(光電変換)を担い、光合成電子伝達の最初の化学反応場である。
2. アンテナ複合体の色素成分と役割
- アンテナ複合体はクロロフィルaに加え、多くのクロロフィルbや**カロテノイド(カロテン、キサントフィルなど)**を含む。
- これらの色素は広い波長の光を吸収し、吸収した光エネルギーを反応中心のクロロフィルaに効率よくエネルギー移動(共鳴エネルギー移動)によって伝えている。
- アンテナ複合体は「光捕集器」として機能し、多くの光子を捕まえて反応中心の光反応効率を高める役割がある。
3. 色素が光合成に関与していることを示す証拠
- 吸収スペクトルの対応
光合成活性のピークはクロロフィルやカロテノイドの吸収波長と一致する。特に、光合成作用スペクトルと色素の吸収スペクトルが重なることが示されている。 - 色素除去・変異による光合成低下
クロロフィルやカロテノイドを化学的に除去、または遺伝子操作で色素合成酵素を欠失させた場合に光合成能が顕著に低下する実験結果。 - 蛍光緩和とエネルギー移動の直接観察
アンテナ複合体から反応中心へのエネルギー移動は蛍光スペクトル解析や時間分解蛍光法で実験的に観測されている。 - 反応中心の電子伝達活性
反応中心クロロフィルaの励起により電子移動が始まることは、光化学活性の電気化学測定や機能解析で確証されている。
このように、反応中心とアンテナ複合体の各色素は役割分担しつつ密接に協力して光エネルギーを利用することで高効率に光合成反応を駆動している。光化学系は反応中心とアンテナ複合体からなり、反応中心には主にクロロフィルaが含まれていて、光吸収により電子を励起し電子伝達の開始点となる。アンテナ複合体にはクロロフィルa、クロロフィルb、カロテノイドが含まれ、広範な波長の光を吸収して反応中心に効率よくエネルギーを伝達する。これら色素が光合成に関与している証拠として、光合成活性と色素の吸収スペクトルの一致、色素除去や合成酵素の変異で光合成能低下が起きること、蛍光スペクトルや時間分解蛍光法でのエネルギー移動の測定、反応中心クロロフィルaの電子伝達活性の検出などがある。
緑色および紅色細菌の光合成ではO2は発生しない。なぜか。これらの生物が光合成を使ってATPを合成できるのはなぜか.これらの生物では何が電子供与体となっているのか
なぜO2が発生しないか
- 緑色細菌や紅色細菌は光化学系を1つしか持たず(光化学系I型またはⅡ型)、水の分解(水酸化)を行わないためO2を発生しない。
- 彼らは「非酸素発生型光合成」を行い、光エネルギーを用いて電子伝達を進めるが、電子供与体として水ではなく、H2S(硫化水素)や有機化合物を用いるため、酸素は生成されない。
なぜATPを合成できるか
- これらの細菌も光化学反応によって膜電位やプロトン勾配(pmf)を作り出し、このpmfを使ってF型ATP合成酵素でATPを合成する。
- 非循環的電子伝達系を持つ場合もあれば、循環的電子伝達によりプロトン駆動力を成立させる場合もあり、いずれも光エネルギーを化学的エネルギー(ATP)に変換できる。
電子供与体
- 主に硫化水素(H2S)、その他の無機硫黄化合物や有機分子が電子供与体となる。
- 例えば、CO2 + 2H2S → [CH2O] + H2O + 2S のように、硫化水素が酸化され、その電子が光合成電子伝達に供給される。
まとめると、
緑色および紅色細菌は水を電子供与体とせず、O2発生を伴わない非酸素発生型光合成を行うが、光エネルギーで膜電位・プロトン勾配を作りATPを合成して生存している。電子供与体はH2Sなどの硫黄化合物や有機分子である。
葉緑体には二つの光化学系が存在する。それぞれの機能は何か。線形電子伝達で、光吸収から NADPH 生成までの電子の流れを図で示せ。NADPHにたくわえられたエネルギーから何がつくられるのか
1. 光化学系I(PSI)と光化学系II(PSII)の機能
- 光化学系II(PSII)
水を分解して電子を取り出し、光エネルギーで励起した電子を電子伝達鎖に渡す。酸素とプロトンも放出される。 - 光化学系I(PSI)
PSIIから電子が伝達されてきた後、さらに光エネルギーを受けて電子を励起し、最終的にNADP+をNADPHに還元する。
2. 線形電子伝達の電子の流れ(図イメージ説明)
水→PSII電子→プラストキノン (PQ)→シトクロムb6f複合体→プラストシアニン (PC)→PSI→光照射電子→フェレドキシン (Fd)→NADP+→フェレドキシン-NADP還元酵素NADPH水PSII電子→プラストキノン (PQ)→シトクロムb6f複合体→プラストシアニン (PC)→PSI光照射電子→フェレドキシン (Fd)→NADP+フェレドキシン-NADP還元酵素NADPH
\[
\text{水} \xrightarrow{PSII} \text{電子} \rightarrow \text{プラストキノン (PQ)}
\]
\[
\rightarrow \text{シトクロムb6f複合体} \rightarrow \text{プラストシアニン (PC)}
\]
\[
\rightarrow \text{PSI} \xrightarrow {光照射} \text{電子} \rightarrow \text{フェレドキシン (Fd)}
\]
\[
\rightarrow \text{NADP}^+ \xrightarrow[\text{フェレドキシン:NADP還元酵素}]{} \textbf{NADPH}
\]
- 水分子がPSIIで酸化され、電子が放出される。
- 電子は順にPQ、シトクロムb6f複合体、PCを経てPSIに渡される。
- PSIで再度光エネルギーを吸収し電子が励起され、フェレドキシンに渡される。
- フェレドキシンNADP還元酵素の作用でNADP+がNADPHに還元される。
3. NADPHに蓄えられたエネルギーの用途
- NADPHは**カルビン・ベンソン回路(暗反応)**でCO2還元のための還元力として使われ、グルコースなどの炭水化物が合成される。
- つまりNADPHは光合成の炭素固定過程における還元エネルギー貯蔵体である。
これにより光合成は、光エネルギーをATPとNADPHの化学エネルギーに変換し、そのエネルギーでCO2を還元して有機物を生み出すシステムとなっています。光化学系II(PSII)は水の分解により電子とプロトンを取り出し、酸素を発生させる。光化学系I(PSI)は高エネルギー電子を受け取って、NADP⁺をNADPHに還元する。
線形電子伝達の電子の流れ(矢印の順に電子移動):
水 → PSII → プラストキノン(PQ)→ シトクロムb6f複合体 → プラストシアニン(PC)→ PSI → フェレドキシン(Fd)→ フェレドキシン-NADP還元酵素 → NADP⁺ → NADPH
NADPHに蓄えられたエネルギーはカルビン・ベンソン回路でのCO2還元に使われ、炭水化物が合成される。
暗所ではCO2を固定するカルビン回路の反応は行われない。その理由として何が考えられるか。これらの反応はどのようにして光によって調節されるのか
カルビン回路が暗所で働かない理由
- カルビン回路は酵素反応の集合体で直に光エネルギーを必要としない「暗反応」と呼ばれるが、実際には光合成の明反応(光化学系)で生成されるATPやNADPHの供給がなければ進まない。暗所ではそれらの供給がなくなるため、カルビン回路はほとんど停止する。
- また、カルビン回路の主要酵素であるRubisCOや他のいくつかの酵素は光により活性化されることが知られている。光が弱い・ないと、これら酵素の活性が下がるため回路が働かなくなる。
光による調節機構
- チオレドキシンシステム(レドックス制御)
光の力で電子伝達系から還元力が伝わり、フェレドキシン-チオレドキシンシステムが還元され、これがカルビン回路の酵素(フルクトースビスホスファターゼ、セドヘプツロースビスホスファターゼ、5-ホスホリブロキナーゼなど)のジスルフィド結合を還元して酵素活性を高める。 - Mg²⁺濃度とpHの変動
光によってチラコイド内にプロトンが蓄積され、ストロマのpHが上昇(アルカリ側へ移動)し、Mg²⁺がストロマに移動してMg²⁺濃度が上昇することも酵素活性化に寄与する。 - RubisCOの活性化にはRubisCO活性化酵素によりATP依存的な反応と、炭酸イオンやMg²⁺の存在が重要。
まとめ
カルビン回路は暗所でも化学的には動きうるが、光合成の明反応で生成されるATP・NADPHやチオレドキシンを介する還元調節に依存し、その供給・活性化が止まれば暗所では機能しなくなる。植物はこのメカニズムにより光環境の変化に応じて効率よくカルビン回路の活性を調節している。
地球上に最も大量に存在するタンパク質であるrubiscoは光合成生物における炭水化物合成に重要な役割を果たす。rubiscoとは何で、細胞内のどこに存在し、どのような機能を果たしているのか
ルビスコとは
ルビスコ(正式名:リブロース-1,5-ビスリン酸カルボキシラーゼ/オキシゲナーゼ、ribulose-1,5-bisphosphate carboxylase/oxygenase)は、光合成の炭素固定を触媒する酵素であり、地球上でもっとも大量に存在するとされるタンパク質である。これは5炭糖のリブロース-1,5-ビスリン酸(RuBP)にCO2を付加し、その結果3-ホスホグリセリン酸(3-PGA)を生成する反応を通じて、無機CO2を有機化合物に変換する。
細胞内の存在場所
- 真核植物では、ルビスコは葉緑体のストロマ(基質領域)に存在し、カルビン回路の中心酵素として機能している。
- 一般に、大きな酵素複合体として、大サブユニット8個と小サブユニット8個からなる複合体構造をとる。
機能
- ルビスコは光合成におけるCO2固定酵素として作用し、CO2分子をRuBPに付加して3-PGAを生じる。この反応は炭素固定の最初の非可逆ステップであり、これにより生物は無機炭素を有機物に変換し、植物体の炭水化物合成を支えている。
- ルビスコは同時に酸素を基質にして酸素化反応も触媒し、この反応は光呼吸として知られる代謝経路につながるが、主な役割はカルビン回路でのCO2固定である。
まとめると、ルビスコは光合成の中心的なCO2固定酵素であり、葉緑体ストロマに存在してカルビン回路の炭素同化を促進し、植物や藻類の有機物合成の基盤となっている。

コメント